






 下载:
下载:


-
全球气候变化下物种分布是当下植物地理学、生态学研究的热点问题[1]。气候变化对全球生物多样性产生了巨大的影响,由气候变化导致的物种栖息环境的破碎化是物种灭绝或濒临灭绝的重要原因[2]。海拔、坡度、坡向是影响区域物种组成重要的地形因子[3],相对于其他环境因子更易于测量[4],它们直接或间接的影响物种的土壤含水率及光照条件等,从而影响物种的分布[5, 6]。气候因子与地形因子是影响物种分布的主要环境因子,通过环境因子对物种的生境现状进行模拟或对过去以及未来的生境进行预测,初步了解其潜在的适生区,对开展生物多样性保护,生态系统可持续发展具有较高的指导意义[7]。
利用物种分布模型模拟物种的潜在适生区是一种了解环境因子对物种分布影响的有效途径[8]。物种分布模型(Species distribution model,SDM),又称生态位模型(Niche model),是评估环境因子对物种分布影响以及重建物种历史分布的主要工具,对于资源管理和物种保护规划均具有重要的意义[9]。它是依托生态位理论,通过收集物种(植物或动物)的分布的实际存在点,以影响物种分布的相关环境因子为基础,探究环境因子对物种分布点的影响,明确其主要的环境因子,最后模拟或预测出物种的潜在分布区,探究物种在生态位空间的实际状况[10]的计量模型[11]。物种分布模型的类型和数量持续不断增加,其中MaxEnt(最大熵模型)被许多学者广泛运用,其原因是因为该模型易于上手操作,建模界面简单直观,且具有强大的预测精度,对于生态位理论的理解更加容易[12]。
黄杉隶属于松科 ( Pinaceae) 黄杉属 ( Pseudotsuga) ,第三纪遗留植物、已被列入国家二级重点保护树种名录,为我国特有种,且是优质的种质资源。当前对黄杉或黄杉属开展研究的主要集中于黄杉的群落结构、群落的海拔梯度格局以及资源利用现状等[13-15]。李望军等[16]利用MaxEnt模拟了气候环境因子影响下贵州省天然黄杉林的潜在分布格局,预测了贵州省天然黄杉林的潜在分布区,明确了主导气候因子,但一方面未考虑在训练模型前对气候因子的相关性及共线性分析,另一方面未考虑将地形因子作为MaxEnt模型的驱动因子,缺乏探究地形因子影响黄杉潜在分布的内在机理。四川省与贵州省整体均为温润气候,但四川省较贵州在地形上具有更大的整体落差,从而可能导致区域小气候分异的形成。此外,四川省是我国黄杉主要分布的省份之一,保护工作还位于初级阶段,仅有文剑然[17]对云南、贵州、四川三省的黄杉森林类型,群落结构、更新动态进行了调查研究,研究结果表明黄杉更新依赖于一定的自然干扰;庞鑫[18]对四川省凉山州西昌黄杉(Pseudotsuga xichangensis)这一极小种群物种开展了种群繁育研究,除此之外对于四川省黄杉植物的研究寥寥无几。对于利用物种分布模型对四川省黄杉这一物种开展潜在适生区模拟研究还存在空白,保护工作的前提是要了解黄杉的潜在适生区分布现状,但考虑到野外实际调研面积大,工作量繁重,单纯凭借野外调研很难以有效初步且迅速了解黄杉在四川省的分布现状,通过物种分布模型来进行大尺度的适生区模拟是目前阶段有效的途径。
因此,利用收集到的黄杉实际分布点数据,基于最大熵模型平台,利用生物气候数据(1970-2020年)以及地形因子作为环境因子来模拟黄杉在四川省的适生区分布格局,探究影响其分布的主要环境因子,明确其潜在适生区分布,旨在为四川省黄杉的野外调查及保护工作的开展提供理论依据。
-
黄杉物种分布点位数据来源于多样性信息生物网络数据库(
https://www.gbif.org/ )和中国数字植物标本馆(http://www.cvh.ac.cn/ )以及部分野外实地调查获取的点位数据。先对获取的数据进行清洗,剔除重复和无效的数据点,得到了20个黄杉野外实际的分布点。 -
(1)气候因子
过去时期(1970-2000)的生物气候数据来源于世界气候数据库WorldClim(
http://www.worldclim.org )。由于世界气候数据库并未提供现在时期(2000-2020)的生物气候数据,但给出了生物气候数据生成的方法,参考利用R语言中的dismo包中的biovars函数(https://www.worldclim.org/data/bioclim.html )可得出逐年的生物气候数据,再利用ArcGIS中的像元统计就可得出(2000-2020)这一时序范围的19个生物气候数据。具体方法为,首先,从国家地球系统科学数据中心(http://www.geodata.cn )获取2000-2018年的月最大、最小气温及降水的栅格数据,然后获取美国国家海洋和大气管理局(https://www.ncei.noaa.gov )2019-2020年的最大、最小气温数据统计资料,利用ANUSPLIN4.36软件中的薄板光滑样条函数(Thin Plate Smoothing Spline Method)插值出2019、2020年的最大最小温度,再利用R语言“dismo”程序包中的“biovars”函数得到2000-2020年逐年的生物气候数据,最后通过ArcGIS的像元统计工具得到(1970-2020)的19个生物气候数据。生物气候数据之间的高度相关性和共线性很容易导致模型的过度拟合,从而影响最终预测的准确性。因此,利用R语言“usdm”程序包筛选出秩相关系数(spearman)小于0.8,方差膨胀因子(Variance inflation factor)小于10的生物气候数据(表1),筛选结果保留Bio2、Bio4、Bio7、Bio9、Bio14、Bio15这6个变量作为气候因子。
表 1 最大熵模型环境因子贡献率与置换重要性及方差膨胀因子值
Table 1. Maxent model factor Percent contribution,Permutation importance and Variance inflation factor value.
环境因子
(Environmental factor)贡献率
(Percent contribution/%)累积贡献率
(Accumulative contribution rate/%)置换重要性
(Permutation importance/%)方差膨胀因子
(Variance inflation factor)BIO7 45.2 45.2 13.3 6.291263 DEM 19.9 65.1 35.5 - BIO4 17.4 82.5 10.1 2.931618 BIO2 7.3 89.8 13.2 1.010199 SLOPE 3.3 93.1 6.5 - BIO15 3 96.1 4.6 1.756841 BIO9 2.6 98.7 16.5 4.352802 ASPECT 1.1 99.8 0.2 - BIO14 0.2 100 0.1 3.613532 (2)地形因子
DEM数据来源于世界气候数据库WorldClim,坡度和坡向数据利用ArcGIS10.8软件从DEM数据中提取。忽略长时间序列地形变化对物种分布的影响[19]。
以上数据的分辨率均为30″×30″。
-
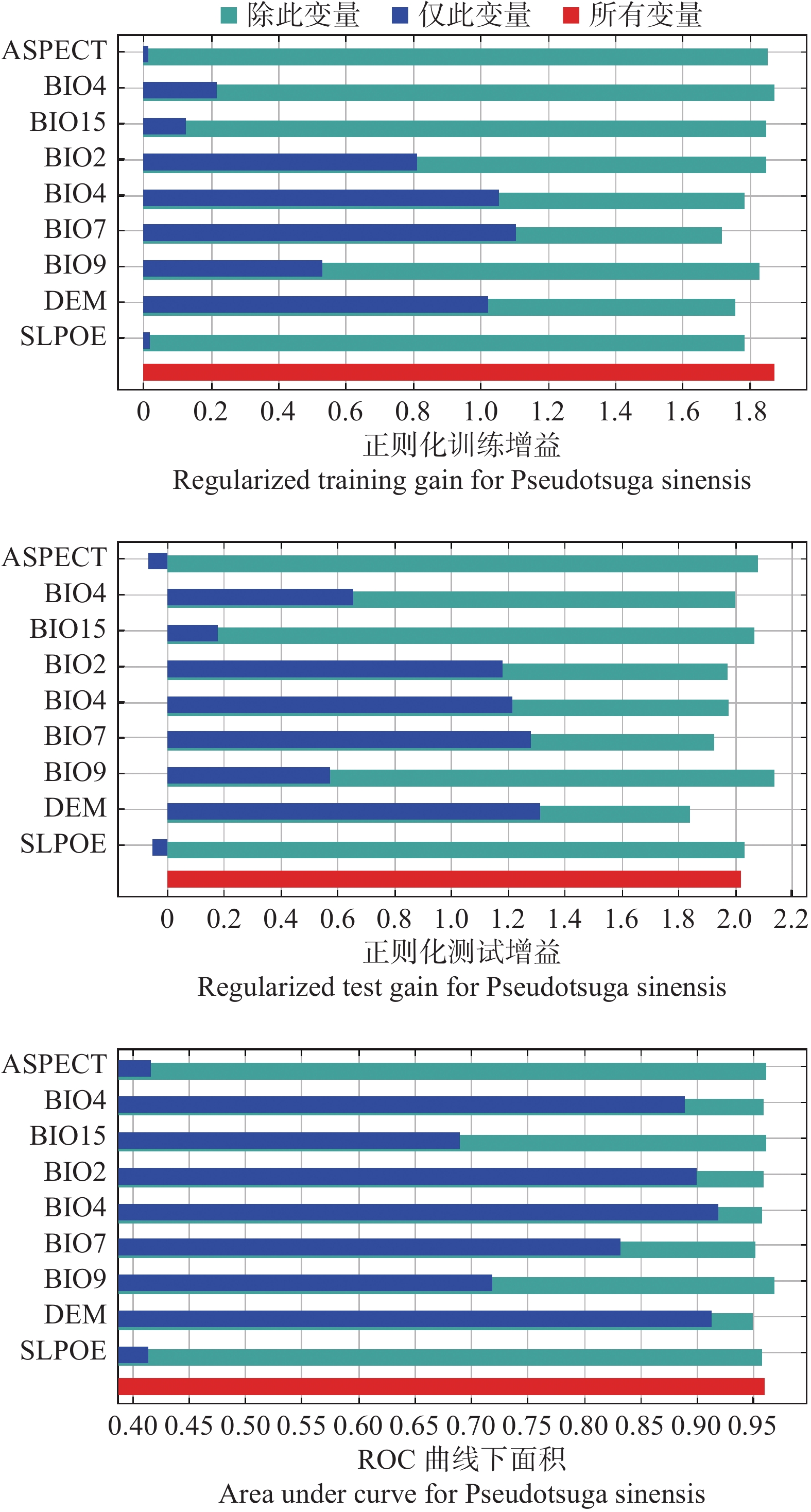
Jackknife测试、贡献率、置换重要性是常用的评估环境因子对模型影响的指标,从而评估不同环境因子对四川省黄杉潜在适生区的重要性[22]。Jackknife测试结果表明(见图1),仅此变量时,正则化训练增益、测试增益、ROC曲线下面积(AUC)显示各环境因子得分值不尽相同,综合三个Jackknife测试结果,DEM为得分值最高的环境因子。贡献率和置换重要性结果(见表1)显示,DEM仍为综合得分最高的环境因子,通过统计环境因子贡献率、置换重要性、仅有此变量时Jackknife测试结果排名(见表2),可以看出最终综合排名第一的环境因子为海拔(DEM)。
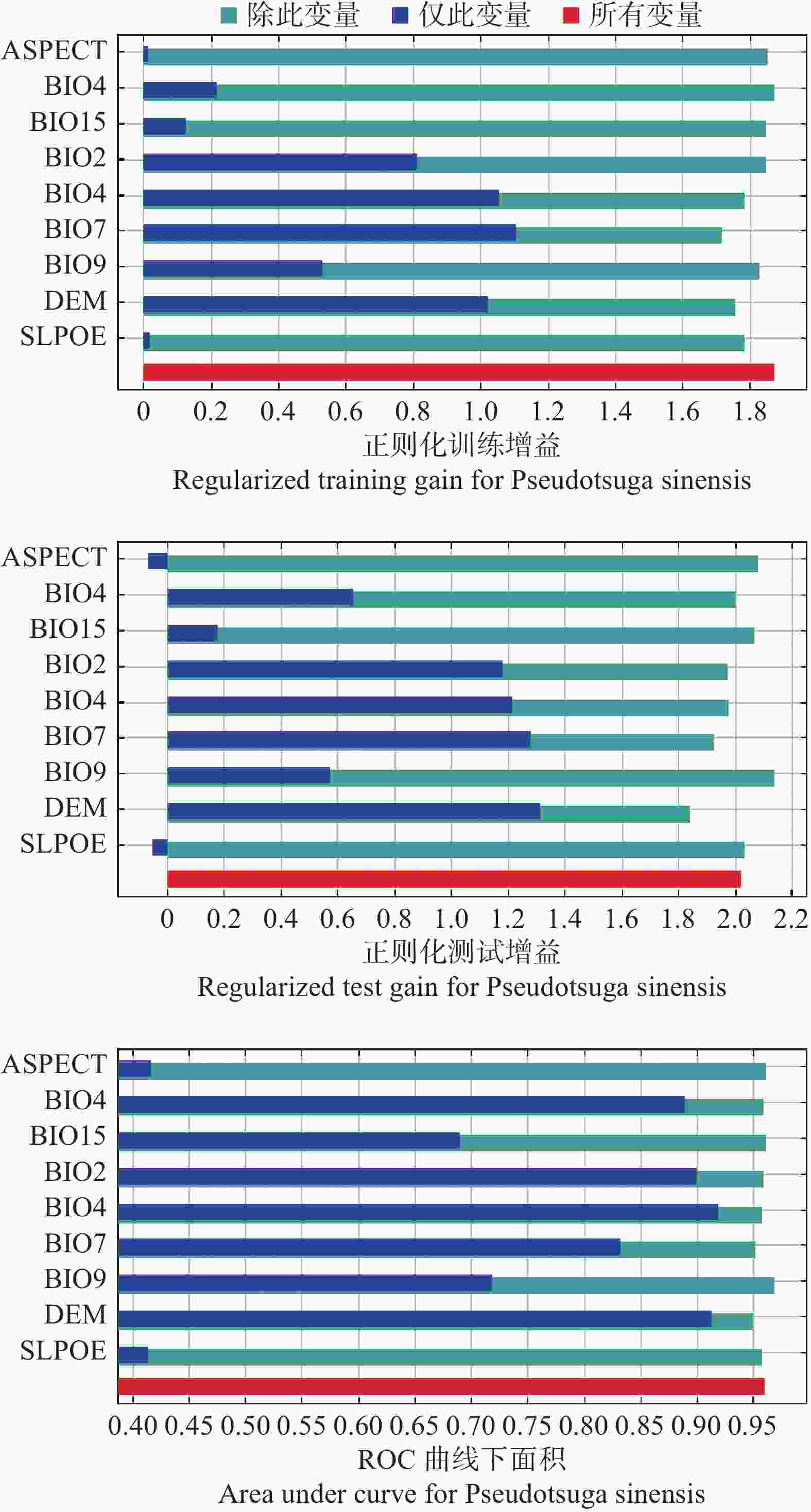
图 1 最大熵模型刀切检验(Jackknife)测试结果
Figure 1. Jackknife test of the MaxEnt model
表 2 贡献率、置换重要性、Jackknife测试排名
Table 2. Percent contribution, Permutation importance and Jackknife test ranking
环境因子
(Environmental factor)因子名称
(Factor name)CP PP JTRAIN JTEST JAUC 综合排名
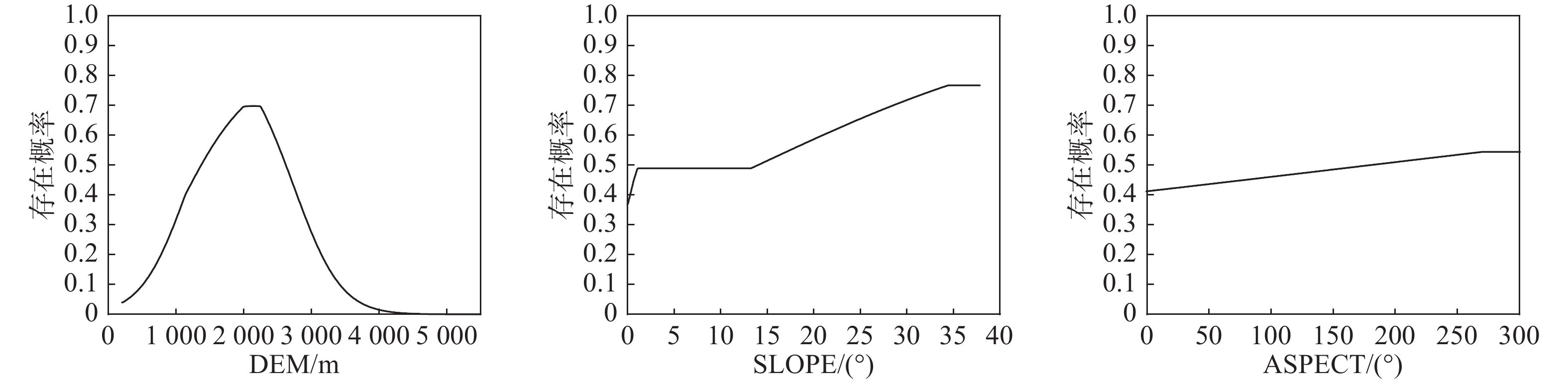
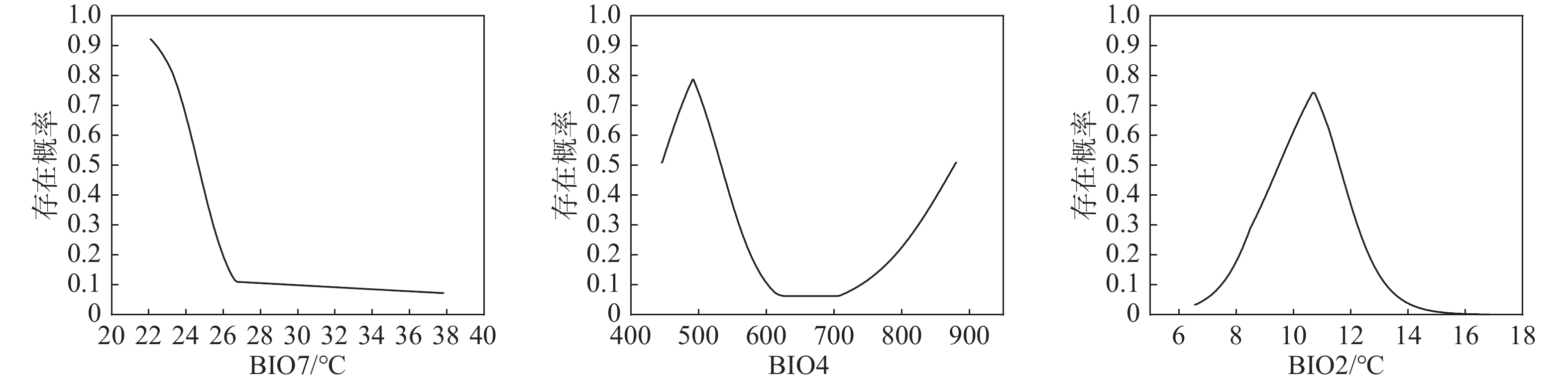
(Integrated ranking)DEM 海拔 2 1 3 1 2 1 BIO7 年最大温差 1 3 1 2 5 2 BIO4 温度季节变化 3 5 2 3 1 3 BIO2 昼夜温差月均值 4 4 4 4 3 4 BIO9 最干季度平均温 7 2 5 6 6 5 BIO15 降水量季节变化 6 7 6 7 7 6 BIO14 最干月降水量 9 9 6 5 4 6 SLOPE 坡度 5 6 8 8 9 7 ASPECT 坡向 8 8 7 9 8 8 注:CP代表贡献率排名,PP代表置换重要性排名,JTRAIN代表训练集Jackknife排名,JTEST代表测试集Jackknife排名,JAUC代表ROC曲线下面积Jackknife排名 通过驱动因子响应曲线(见图2和图3),以存在概率大于0.5作为四川省黄杉适生区的判断条件(大于0.5为适宜,小于则不适宜)[22],得到地形因子中海拔的适宜范围约为1375.3 m—2613.7 m,坡度适宜值为大于14.064°,坡向适宜范围为181.202°—269.560°;气候因子中年最大温差的适宜值应小于24.64℃,温度季节性变化较大时较适宜其生存,昼夜温差月均值适宜范围为9.510℃—11.581℃。
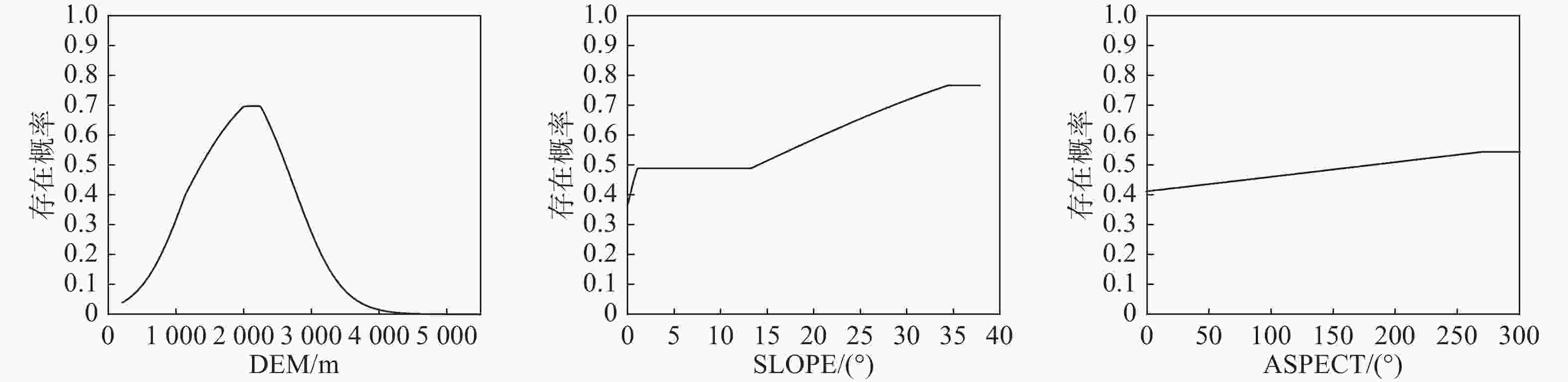
图 2 地形因子响应曲线
Figure 2. Terrain factor response curves

图 3 气候因子响应曲线
Figure 3. Climate factor response curves
-
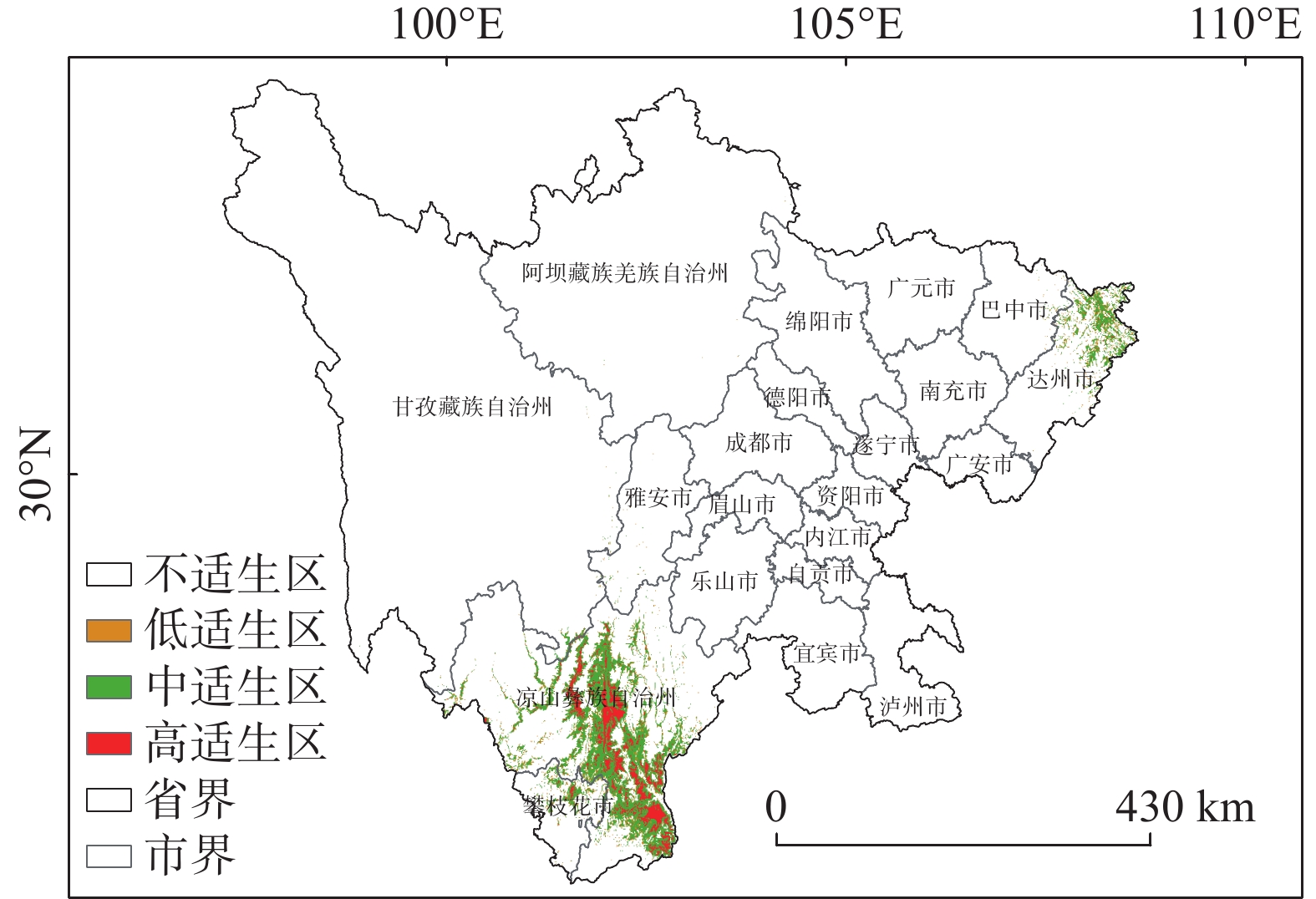
图4的适生区分布图可以看出,适生区总体生境呈现破碎化,适生区主要分布于四川省的部分南部地区,以及部分东部地区,中低适生区分布于高适生区的周围。高适生区主要位于凉山州的中部县区,冕宁县、德昌县、普格县、会理县和会东县等,攀枝花的米易县等,达州市的东北部有零散分布的高适生区。统计结果显示(见表3和图5),适生区总面积为2×104 km2,占研究区总面积的4.12%。低适生区面积为0.273×104 km2,中适生区面积为1.346×104 km2,高适生区面积为0.381×104 km2,分别占适生区总面积的13.7%、67.3%、19%。总适生区面积最大的为凉山彝族自治州,达州市次之;高适生区面积最大的是亦是凉山,高适生区面积为0.367×104 km2,大致沿凉山州境内安宁河谷流域,以及凉山州东南部的黑水河流域分布,海拔跨度区间主要为1000—3500 m,达州市高适生区总面积0.006×104 km2,海拔跨度区间大致为500—1500 m,达州市的高适生区面积略低于攀枝花市,攀枝花市高适生区海拔区间主要为2500—3000 m,高适生区总面积0.007×104 km2。四川省内不适生区海拔跨度为202—6945 m,平均海拔为2624 m;低适生区海拔跨度427—3528 m,平均海拔为1934 m;中适生区海拔跨度424—3626 m,平均海拔2023 m,高适生区海拔跨度688-3274 m,平均海拔2051 m。根据表4的四川省高适生区海拔统计结果显示,高适宜生境主要集中于1500—2000 m,2000—2500 m这两个海拔区间,分别占高适生区总面积的33.3%和48.9%。综合之前的结果,我们可以得知高适生区主要位于凉山彝族自治州,且集中分布在1500—2500 m这一海拔区间。
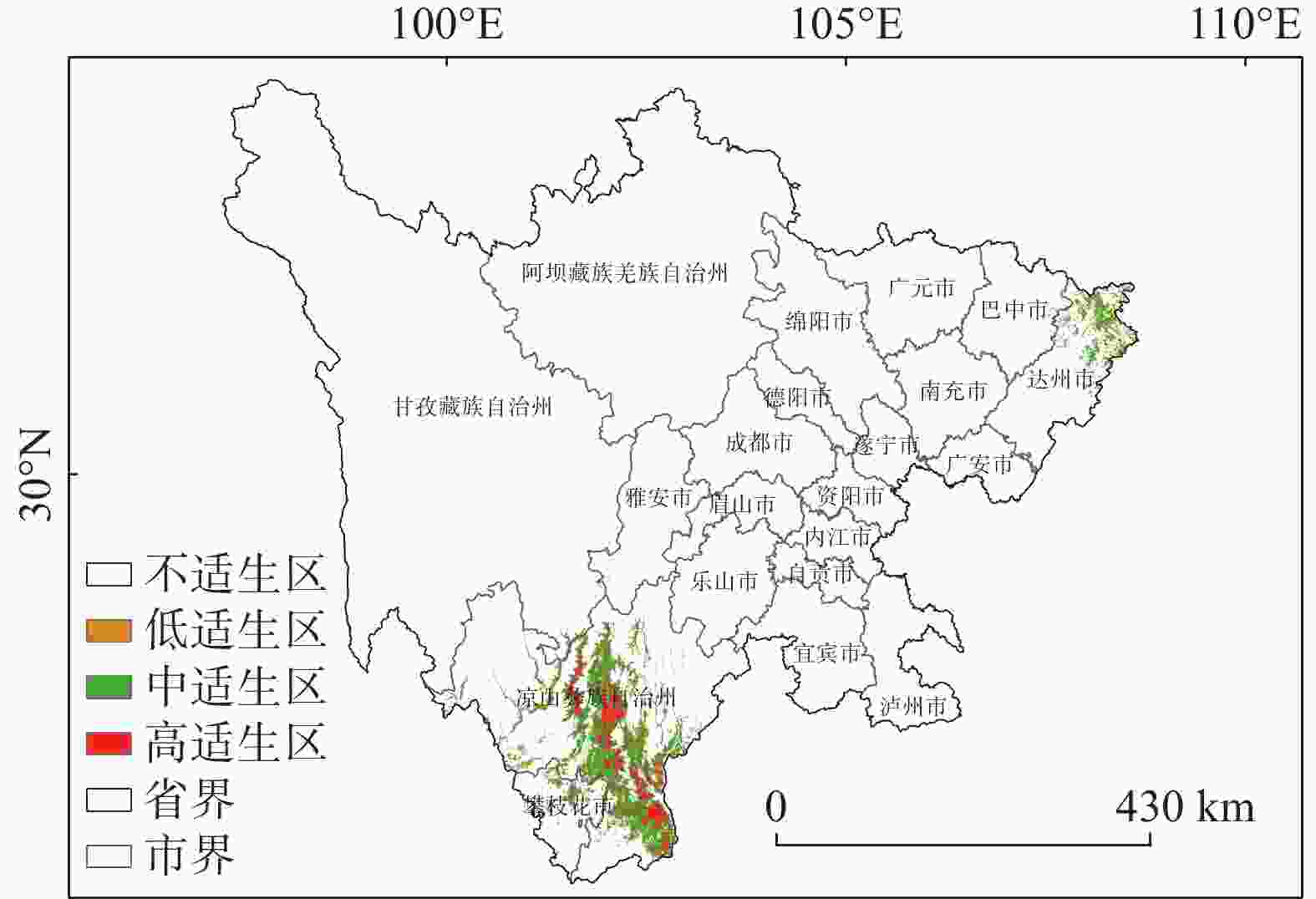
图 4 四川省黄杉适生区分布图
Figure 4. The potential distribution pattern of Pseudotsuga sinensis
表 3 各市州各等级适生区面积统计/×104km2
Table 3. Area statistics of suitable areas of various grades in cities and prefectures
行政区名称
(Administrative district name)不适生
(Unsuitable
area)低适生
(Low suitable
area)中适生
(Moderate suitable
area)高适生
(High suitable
area)总适生区面积
(Total area of
suitable area)凉山彝族自治州 4.251 0.175 1.080 0.367 1.622 达州市 1.402 0.065 0.173 0.006 0.244 攀枝花市 0.605 0.023 0.069 0.007 0.098 甘孜藏族自治州 14.998 0.004 0.019 0.001 0.025 巴中市 1.237 0.003 0.004 0.000 0.007 雅安市 1.500 0.002 0.002 0.000 0.004 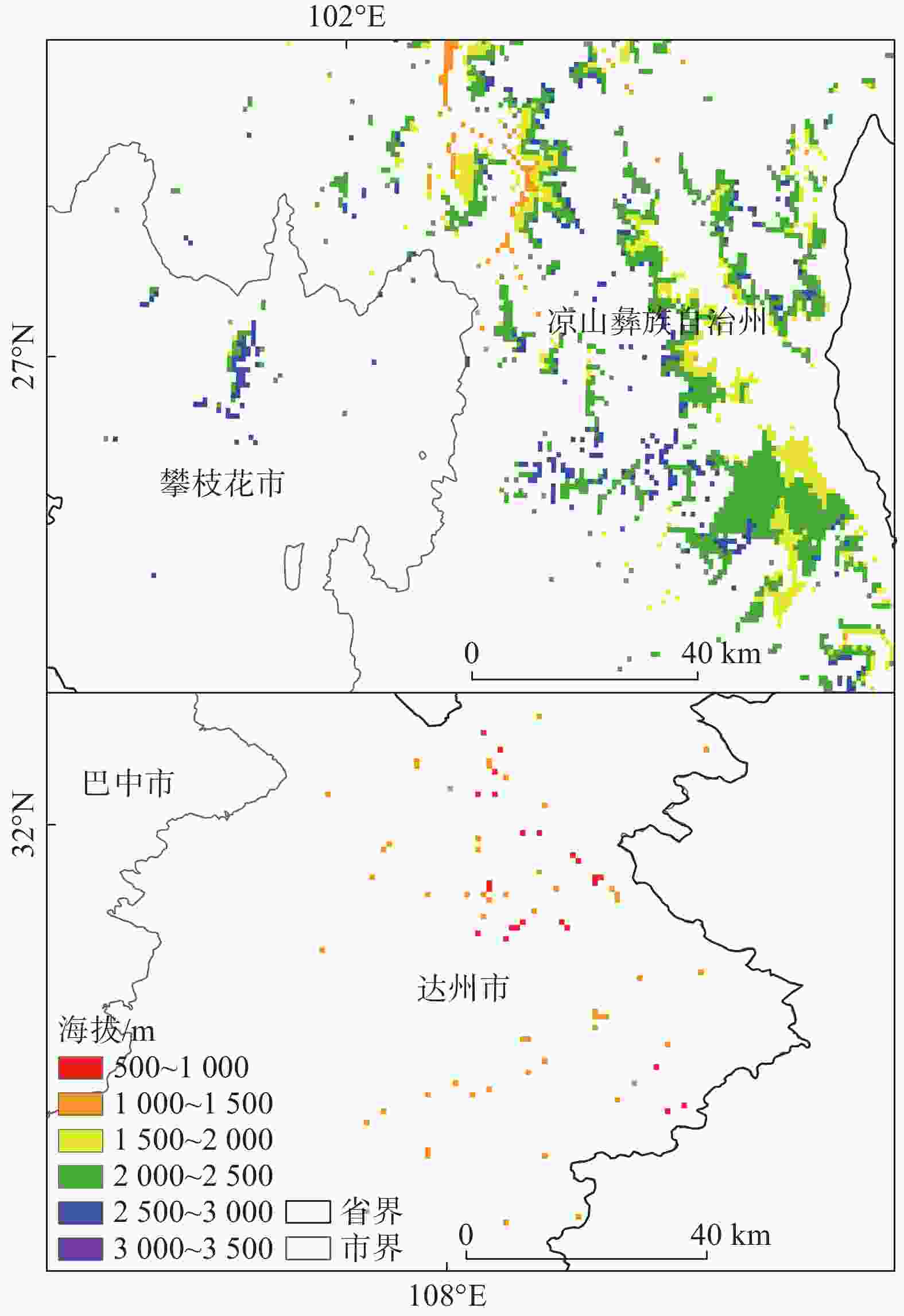
图 5 高适生区海拔分布图
Figure 5. Elevation distribution of high suitable area
表 4 高适生区海拔统计
Table 4. Elevation statistics of high suitable area
海拔(m) 高适生面积
(×104km2)占高适生区
比例(%)占总适生区
比例(%)500-1000 0.002 0.005 0.001 1000-1500 0.027 0.070 0.013 1500-2000 0.127 0.333 0.063 2000-2500 0.186 0.489 0.093 2500-3000 0.038 0.099 0.019 3000-3500 0.002 0.004 0.001 -
模型的性能是通过计算受试者特征曲线下面积(Area Under the Receiver Operator Curve,AUC)来进行评估。一般认为,AUC<0.7代表模型精度低,当AUC处于0.7—0.9之间时表示可采用预测结果,AUC>0.9表示预测结果非常准确[23]。模型结果显示,AUC值为0.988,模拟结果非常准确。
Jackknife测试、贡献率、置换重要性综合排名结果显示,环境因子的综合排序顺序为海拔、温度因子、降水因子、坡度坡向。对植被类型分布格局形成的解释主要都集中在地形、气候等要素,地形因子中,海拔、坡度、坡向作为最重要的地理变量,主要是通过影响土壤温度和水分从而影响植物的分布。海拔是对四川省黄杉植物分布影响最大的因子,海拔适宜范围属于中低海拔地区,该海拔范围内气候适宜,土壤类型多样性趋于平稳[24],是四川省黄杉在此海拔区间内适宜生长的原因之一。海拔响应曲线呈“先促进后抑制”的规律,这与现有的的研究结果高度相似[25],说明过高或过低海拔都不适宜黄杉生长。研究结果表明坡度的适宜范围为大于14.064°,坡向大致为西南坡,坡度与坡向分别影响着土壤的含水率与植被的光照条件,有研究表明,黄杉具有喜光特性,且有适应干旱气候的特性,幼苗相比其他阔叶树种处于劣势,只有在土壤较为干旱的山坡和山脊处,其他树种不适宜生存的条件下,才能发挥其耐干旱优势[26],坡度增加不利于土壤和水分的积累[27],这也印证了黄杉具有一定的耐旱特征。该研究中海拔对黄杉植被的影响明显大于坡度和坡向,是因为不同海拔梯度导致的水热条件不同,海拔可以通过改变水热条件影响黄杉植物的分布。四川省黄杉植物各等级适生区平均海拔差异不明显,但适生区海拔跨度较大,为427 m—3247 m,意味着黄杉具有较为宽广的生态适宜幅度。高适生区海拔统计结果显示,1500—2500 m为主要的高适生区海拔区间,这与李望军在贵州省的研究结果一致[16],该海拔区域内黄杉群落的物种多样性最高,生态系统稳定性较好。黄杉喜温暖湿润,500 m—1500 m这一海拔区间亦为四川省的中低海拔地区,水热条件较为充沛,却大多不是黄杉的高适生区,这可能需结合其生长发育特性,人为干扰,历史演替等因素综合分析。气候因子中温度因子的影响总体大于降水因子,表明四川黄杉植物对于温度的响应程度比降水更加敏感。
黄杉作为国家二级保护植物,除受环境因子影响之外,野外调查时发现受人为砍伐破坏严重,这也是导致黄杉生境破碎的原因之一,后续研究将考虑将人类活动作为驱动因子,能更加准确的模拟出黄杉的潜在适生区;其次,缺乏在气候变化下不同纬度带上黄杉的动态变化探究,因此下一步可考虑将未来气候因素加入进来,模拟未来气候变化下四川省黄杉植物的适生区与变迁情况;最后物种入侵或许是黄杉植物的另一个威胁,通过物种间竞争会减小黄杉幼苗的更新能力,如凉山州危害最严重的入侵植物紫茎泽兰[28],故对黄杉的野外救助工作开展势在必行。
Simulation of potential suitable areas and exploration of dominant environmental factors of Pseudotsuga sinensis in Sichuan Province based on MaxEnt model
More Information-
摘要: 基于四川省黄杉 ( Pseudotsuga sinensis) 分布点数据以及气候(19个生物气候因子,时间跨度为1970—2020)和地形环境因子(海拔、坡度、坡向),通过MaxEnt模型来构建四川省黄杉潜在适生区的生态位模型,探究了影响其分布的主导环境因子,模拟了四川省黄杉植物的潜在分布区。研究结果表明;(1)模型总体的预测精度达到非常精确水平,AUC值达0.988;(2)影响其潜在分布的主要环境因子为海拔(DEM),主导因子的适生区海拔范围为1285—2678.2 m;(3)四川省黄杉高适生区主要位于1500—2500 m这一海拔区间;(4)四川省黄杉潜在适生区面积共2×104 km2,其中高适生区面积0.381×104 km2,中适生区面积1.346×104 km2,低适生区面积0.273×104 km2。综合模型结果我们发现,适生区总体生境呈现破碎化,适生区主要分布于四川省的部分南部地区,以及部分东部地区,中低适生区分布于高适生区的周围,总适生区面积前三为,凉山州、达州市、攀枝花市,高适生区主要位于凉山州的中部县区,冕宁县、德昌县、普格县、会理县和会东县等,攀枝花的米易县等,达州市的东北部有零散分布的高适生区;四川省黄杉潜在适生区的驱动因子中海拔因子的影响大于温度和降水因子,明显大于坡度和坡向,海拔的适生区范围多为中海拔地区,具有较为宽广的生态适宜幅度和一定的耐旱特征。研究结果能有效初步且迅速了解黄杉在四川省的分布现状,对于野外保护工作的开展具有一定的指导意义。Abstract: Based on Pseudotsuga sinensis distribution data,climatic factors(19 biocrimatous factors ,spanning1970-2020) ) and topographic factors (altitude, slope and slope direction) from Sichuan province. The MaxEnt model was used to build a niche model to simulate the potential suitable area of it in Sichuan Province, to explore the dominant environmental factors affecting its distribution, and to simulate the potential distribution area.The results show that; (1) The overall prediction accuracy of the model reached a very accurate level, and the AUC value reached 0.988; (2) The main environmental factor affecting their potential distribution was elevation (DEM), and the habitats of leading factors ranged from 1285 m to 2678.2 m. (3) The high suitable area in Sichuan Province is mainly located in the altitude from 1500 to 2500 m; (4) The potential suitability area of it is 2×104 km2, among which the high suitability area is 0.381×104km2, the middle suitability area is 1.346×104km2 and the low suitability area is 0.273×104km2.Based on the comprehensive results, we found that the overall habitat of the suitable areas was fragmented, and the suitable areas were mainly distributed in some southern areas and some eastern areas of Sichuan Province, and the middle and low suitable areas were distributed around the high suitable areas. The top three suitable areas were Liangshan Prefecture, Dazhou City and Panzhihua City, and the high suitable areas were mainly located in the central counties of Liangshan Prefecture., in Mianning County, Dechang County, Puge County, Huili County and Huidong County, Miyi County of Panzhihua County and the northeast of Dazhou City, there are scattered high suitability areas; Among the driving factors of the potential suitable areas, the influence of altitude factors is greater than that of temperature and precipitation factors, and significantly greater than that of slope and aspect. Most of the suitable areas of altitude are middle-altitude areas, which have relatively broad ecological suitability range and certain drought tolerance characteristics.The results of this study can effectively preliminarily and rapidly understand the distribution status of it in Sichuan Province, and have certain guiding significance for the development of field conservation work.
-
Key words:
- Pseudotsuga sinensis;
- MaxEnt model;
- potential distribution area;
-
表 1 最大熵模型环境因子贡献率与置换重要性及方差膨胀因子值
Tab. 1 Maxent model factor Percent contribution,Permutation importance and Variance inflation factor value.
环境因子
(Environmental factor)贡献率
(Percent contribution/%)累积贡献率
(Accumulative contribution rate/%)置换重要性
(Permutation importance/%)方差膨胀因子
(Variance inflation factor)BIO7 45.2 45.2 13.3 6.291263 DEM 19.9 65.1 35.5 - BIO4 17.4 82.5 10.1 2.931618 BIO2 7.3 89.8 13.2 1.010199 SLOPE 3.3 93.1 6.5 - BIO15 3 96.1 4.6 1.756841 BIO9 2.6 98.7 16.5 4.352802 ASPECT 1.1 99.8 0.2 - BIO14 0.2 100 0.1 3.613532  下载: 导出CSV
下载: 导出CSV
表 2 贡献率、置换重要性、Jackknife测试排名
Tab. 2 Percent contribution, Permutation importance and Jackknife test ranking
环境因子
(Environmental factor)因子名称
(Factor name)CP PP JTRAIN JTEST JAUC 综合排名
(Integrated ranking)DEM 海拔 2 1 3 1 2 1 BIO7 年最大温差 1 3 1 2 5 2 BIO4 温度季节变化 3 5 2 3 1 3 BIO2 昼夜温差月均值 4 4 4 4 3 4 BIO9 最干季度平均温 7 2 5 6 6 5 BIO15 降水量季节变化 6 7 6 7 7 6 BIO14 最干月降水量 9 9 6 5 4 6 SLOPE 坡度 5 6 8 8 9 7 ASPECT 坡向 8 8 7 9 8 8 注:CP代表贡献率排名,PP代表置换重要性排名,JTRAIN代表训练集Jackknife排名,JTEST代表测试集Jackknife排名,JAUC代表ROC曲线下面积Jackknife排名
下载: 导出CSV
表 3 各市州各等级适生区面积统计/×104km2
Tab. 3 Area statistics of suitable areas of various grades in cities and prefectures
行政区名称
(Administrative district name)不适生
(Unsuitable
area)低适生
(Low suitable
area)中适生
(Moderate suitable
area)高适生
(High suitable
area)总适生区面积
(Total area of
suitable area)凉山彝族自治州 4.251 0.175 1.080 0.367 1.622 达州市 1.402 0.065 0.173 0.006 0.244 攀枝花市 0.605 0.023 0.069 0.007 0.098 甘孜藏族自治州 14.998 0.004 0.019 0.001 0.025 巴中市 1.237 0.003 0.004 0.000 0.007 雅安市 1.500 0.002 0.002 0.000 0.004
下载: 导出CSV
表 4 高适生区海拔统计
Tab. 4 Elevation statistics of high suitable area
海拔(m) 高适生面积
(×104km2)占高适生区
比例(%)占总适生区
比例(%)500-1000 0.002 0.005 0.001 1000-1500 0.027 0.070 0.013 1500-2000 0.127 0.333 0.063 2000-2500 0.186 0.489 0.093 2500-3000 0.038 0.099 0.019 3000-3500 0.002 0.004 0.001
下载: 导出CSV
-
[1] 孟影,马姜明,王永琪,等. 基于Maxent模型的檵木分布格局模拟[J]. 生态学报,2020,40(22):8287−8296. [2] Keinath D A, Doak D F, Hodges K E, et al. A global analysis of traits predicting species sensitivity to habitat fragmentation.[J]. Global Ecology & Biogeography, 2017(No.1): 115−127. [3] 左永令,杨小波,李东海,等. 环境因子对海南岛野生兰科植物物种组成与分布格局的影响[J]. 植物生态学报,2021,45(12):9. [4] 沈泽昊,胡志伟,赵俊,等. 安徽牯牛降的植物多样性垂直分布特征——兼论山顶效应的影响[J]. 山地学报,2007,25(2):9. [5] 张子良,于飞,王得祥. 环境因子对秦岭3种典型次生林群落物种组成的影响[J]. 西北农林科技大学学报:自然科学版,2014,42(08):69−78. [6] 陈瑶,胥晓,张德然,等. 四川龙门山西北部植被分布与地形因子的相关性[J]. 生态学杂志,2006,25(9):1052−1055. [7] 油志远,王茗,鲁碧耕,等. 基于MaxEnt模型预测蓝马鸡的潜在分布区[J]. 生态学杂志,2022,41(11):2271−2277. [8] Zimmermann N E, Edwards T C, Graham C H, et al. New trends in species distribution modelling[J]. ECOGRAPHY, 2010(No.6): 985−989. [9] Antoine Guisan N E Z. Predictive habitat distribution models in ecology[J]. Ecological Modelling, 2000(No.2-3): 147−186. [10] Elith J, Leathwick J R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. [J]. Annual Review of Ecology, Evolution & Systematics, 2009(No. 1): 677-697. [11] 乔慧捷,胡军华,黄继红. 生态位模型的理论基础、发展方向与挑战[J]. 中国科学:生命科学,2013,43(11):915−927. [12] Wang T, Zhang Z, Liu L, et al. Simulation of the Potential Distribution of the Glacier Based on Maximum Entropy Model in the Tianshan Mountains, China[J]. Water, 2021(No.1541): 1541): 1541. [13] 孙卫邦,孔繁才,周元,等. 黄杉属植物在云南的资源现状及保护利用研究[J]. 广西植物,2003,23(1):4. [14] 何斌,李青,陈群利,等. 黔西北黄杉群落物种多样性的海拔梯度格局[J]. 生态环境学报,2021,30(6):10. [15] 陈文红,税玉民,王文,等. 云南易门翠柏和黄彬的群落调查及保护[J]. 云南植物研究,2001(第2期):189−200. [16] 李望军,冯图,周瑞伍,等. 基于Maxent模型的贵州省天然黄杉林的潜在分布预测研究[J]. 西部林业科学,2019,48(03):47−52. [17] 文剑然. 中国西南地区孑遗植物黄杉的森林类型、群落结构与更新动态[D]. 云南大学,2022. [18] 庞鑫. 珍贵观赏植物西昌黄杉种群特征及保护利用策略研究[D]. 成都理工大学,2023. [19] 廖剑锋,易自力,李世成,等. 基于Maxent模型的双药芒不同时期潜在分布研究[J]. 生态学报,2020,40(22):9. [20] 赵管乐,刘勤,彭培好. 基于Sentinel2影像与MaxEnt模型识别云南华坪县芒果种植区[J]. 中国农业科技导报,2022,24(3):111−119. [21] 赵光华,樊保国. 末次间冰期以来濒危植物藤枣适生区空间迁移预测[J]. 西南农业学报,2021(第1期):174−182. [22] 唐自豪,刘贤安,彭培好,等. 末次冰盛期以来香果树潜在地理分布格局变迁[J]. 生态学报,2023,43(08):3339−3347. [23] 王晓帆,段雨萱,金露露,等. 基于优化的MaxEnt模型预测中国高山栎组植物的历史、现状与未来分布变化[J]. 生态学报,2023(16):1−15. [24] 管磊,王华军,王玉宽,等. 基于GIS的四川省植被类型分布与地质背景的关系研究[J]. 物探化探计算技术,2015,37(5):6. [25] 陈冰瑞,邹慧,孟祥红,等. 气候变化下中国地域内柴胡与狭叶柴胡适生区的分布格局与变迁预测[J]. 生态学报,2022(20):1−13. [26] 陈文红,王文. 云南易门翠柏和黄彬的群落调查及保护[J]. 植物分类与资源学报,2001,23(2):189−200. [27] 李文博,石君杰,张向阳,等. 河南主要山区栎类天然次生林幼树更新特征及影响因子[J]. 河南林业科技,2023,43(02):1−5. [28] 刘海,杜如万,王勇,等. 紫茎泽兰对四川省凉山州共生植物种间联结性及稳定性的影响[J]. 生态学报,2017,37(15):8. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 21
- HTML全文浏览量: 2
- PDF下载量: 4
- 被引次数: 0









