









-
大熊猫(Ailuropoda melanoleuca)是全球生物多样性保护的旗舰物种与伞护物种。过去几十年大熊猫保护取得了举世瞩目的成就,但该物种仍然面临繁殖力弱、存活率低、栖息地破碎化等问题[1]。根据全国第四次大熊猫调查的结果,仍有30%的大熊猫栖息地质量有问题[2]。由于自然隔离和人为干扰,大熊猫栖息地被隔离成33个斑块,面积小于10 000 hm2的斑块有9个,栖息地破碎化将给野生大熊猫种群的生存、繁衍带来威胁[3]。大相岭山系有大熊猫38只[2],是六大山系中种群数量最少的[4]。该区域栖息地面积小、破碎化程度高、人类活动干扰严重,亟待加强保护[4]。伴随大熊猫国家公园的建立,栖息地恢复工作是未来大熊猫国家公园保护管理工作的重点。
大熊猫是森林类型的物种,竹类资源、森林起源以及遮蔽条件是影响大熊猫生境利用的主要因素[5]。森林中的乔木为大熊猫提供躲避、休息以及巢穴。林下竹子作为大熊猫主要的食物来源,是大熊猫生存的基础。已有研究表明,森林群落乔木层郁闭度对林下植物的生长起重要作用[6-9],乔木是森林群落恢复过程中重要的生物因子[10],乔木层通过对光的透射、反射和吸收,直接影响林下光照的强度和分布,而光照对于绝大多数森林树种而言,是决定更新个体能否生存和生长的关键[11]。乔木层郁闭度对竹子的高度、基径和密度等生长发育具有重要的影响[7,12]。因此,研究乔木层结构特征与林下竹子生长指标之间的关系是栖息地恢复的基础,对提升大熊猫栖息地质量、保护大熊猫及全球生物多样性具有至关重要的作用。
八月竹是非常典型的大熊猫取食竹,广泛分布于大相岭山系及其以南的大熊猫栖息地,也是大相岭山系大熊猫最主要的取食来源。基于此,选择大相岭山系的大熊猫栖息地作为研究对象,针对大熊猫栖息地森林群落乔木层结构特征与林下八月竹生长指标开展调查,定量分析两者之间的关系,并建立大熊猫栖息地森林群落乔木层特征与林下八月竹生长关系的模型,以期为该区域大熊猫栖息地恢复提供量化依据。
-
研究区选择在大熊猫国家公园荥经片区羊叉岗旧苗圃基地和码棚头原生森林生态系统林下分布有八月竹的区域(图1)。该区域海拔 2 025~2 276 m,属亚热带高山山地气候。植被类型以冷杉(Abies fabri)、云杉(Picea asperata)、槭树(Acer spp.)、中华木荷(Schima sinensis)、刺榛(Corylus ferox)、荚蒾(Viburnum dilatatum)、水榆花楸(Sorbus alnifolia)、灯笼树(Enkianthus chinensis)、天师栗(Aesculus chinensis)等为主要树种的针阔混交林,林下以八月竹为主,是理想的大熊猫栖息地。
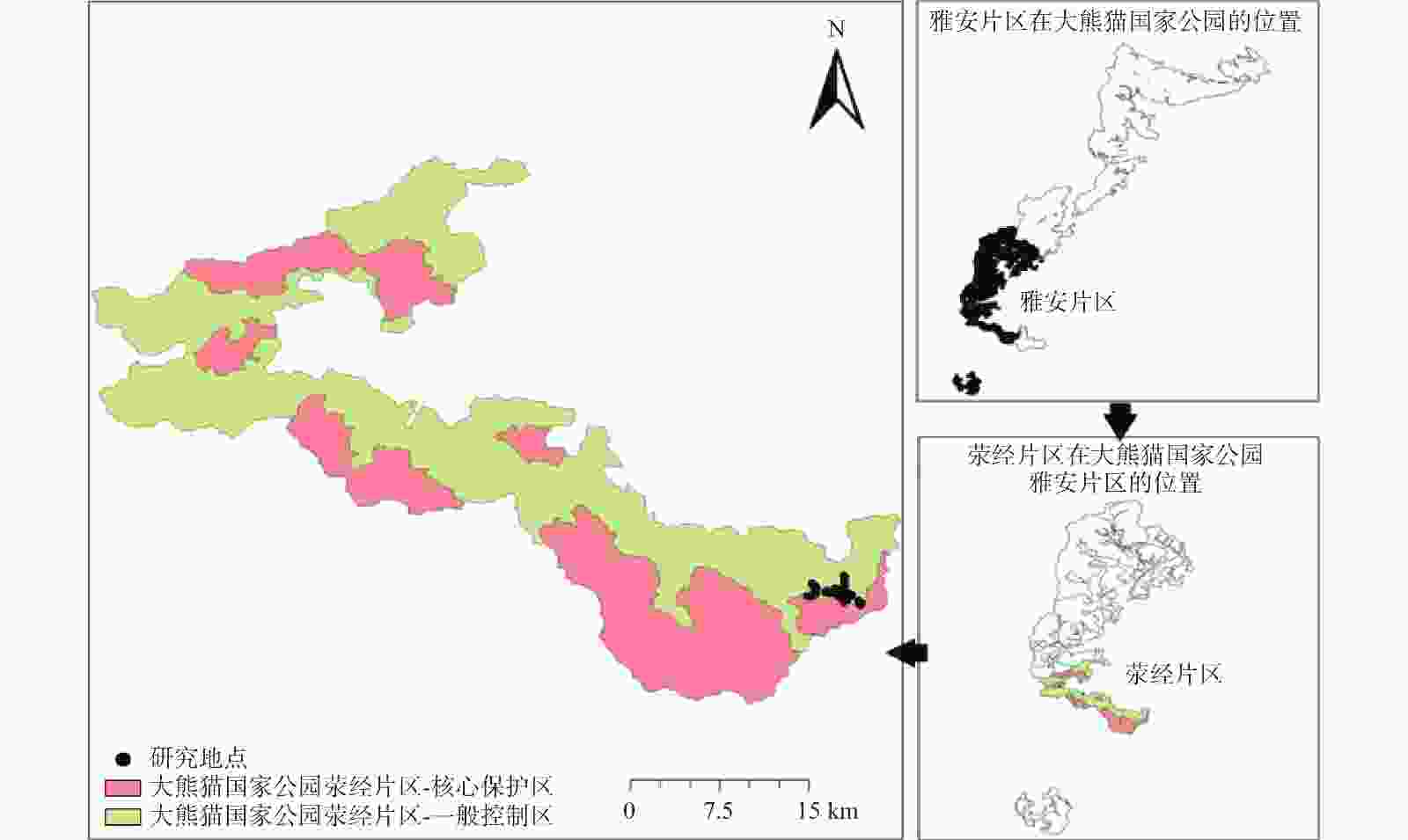
Figure 1. Yingjing district and research area of Giant Panda National Park
-
调查于2022年9月进行。设置3条样线,在样线上随机布设20 m×20 m的乔木样方共79个,记录样方的海拔(m)、坡度(°)、坡向(°)、坡位(背坡、山肩、山顶、麓坡、趾坡)、坡型(均匀坡、复合坡、凹坡、凸坡),以及乔木层郁闭度、乔木株数和乔木平均高度等。每个乔木样方内分别按“品”字形获取3个2 m×2 m的八月竹样方,共计237个八月竹样方。计数每个样方中八月竹发笋量、活竹株数和死竹株数,测不同年份八月竹的基径和高度、笋基径和笋高度。
-
以野外调查所得的237个八月竹样方的样本数据为建模样本。依据乔木层数(3层),将乔木平均高度划分为3个等级:矮:≤6 m,中:7~12 m,高:12 m以上;依据不同样方内的乔木株数之间的差异,将乔木株数划分为3个等级:A:0~15株,B:16~30株,C:31株以上;由于竹子在不同乔木层郁闭度下有差异,而通常乔木层郁闭度又被分为高、中和低3个等级,所以将乔木层郁闭度划分为3个等级:低:0~0.35,中:0.36~0.7,高:>0.7。
利用R软件进行单因素方差分析,以检验乔木层结构特征对八月竹生长指标的影响,然后进行逐步回归分析,选取显著性最高的因子,建立估测模型,根据模型评价指标,选择最优回归模型。对于不同年份八月竹高度和基径、笋高度和基径数据,考虑环境因子(海拔、坡度、坡向、坡位、坡型、植被起源)的随机效应,构建线性混合效应模型[13]:
$ {Y}_{ij}={a}_{0}+{a}_{1}{X}_{ij}+{b}_{i}+{e}_{ij} $ ,其中,$ {Y}_{ij} $ 表示第i类的第j个观测的反应变量,$ {a}_{0} $ 表示固定截距,$ {a}_{1} $ 表示固定斜率,$ {X}_{ij} $ 表示固定效应,$ {b}_{i} $ 表示随机效应(只影响截距),$ {e}_{ij} $ 表示残差;对于八月竹无性系种群数量(包括发笋量、活竹株数和死竹株数)数据,选择泊松分布形式,考虑环境因子(海拔、坡度、坡向、坡位、坡型、植被起源)的随机效应,构建广义线性混合模型[14-16]:$ g\left({\mu }_{ij}\right)={\eta }_{ij}={X}_{ij}\beta +{Z}_{ij}{\mu }_{i} $ ,其中,$ g\left(.\right) $ 表示联结函数,$ {\eta }_{ij} $ 表示条件线性预测值,$ {\mu }_{ij} $ 表示条件均数(考虑了海拔、坡度、坡向、坡位、坡型、植被起源的随机效应),$ {X}_{ij} $ 表示解释变量,$ \beta $ 表示固定效应参数矢量,$ {Z}_{ij} $ 表示与随机效应相关的解释变量,$ {\mu }_{i} $ 表示随机效应。 -
结果显示(见表1):每个八月竹样方内八月竹平均发笋量为(4.89±2.98)株,死竹平均株数为(20.24±7.40)株,活竹平均株数为(10.83±5.96)株,一年生八月竹平均基径为(15.58±3.86)mm,一年生八月竹平均高度为(343.12±112.49)cm,多年生八月竹平均基径为(17.37±2.44)mm,多年生八月竹平均高度为(434.13±942.77)cm,笋平均基径为(20.00±3.53)mm,笋平均高度为(89.88±89.88)cm。每个植物样方内乔木平均高度为(10.47±3.17)m,乔木平均株数为(21.28±15.35)株,乔木平均郁闭度为(0.59±0.16)。
乔木平均高度/m 乔木株数/株 乔木层郁闭度 发笋量/株 死竹株数/株 活竹株数/株 10.47±3.17 21.28±15.35 0.59±0.16 4.89±2.98 3.54±2.26 10.83±5.96 一年生竹基径/mm 一年生竹高度/cm 多年生竹基径/mm 多年生竹高度/cm 笋基径/mm 笋高度/cm 15.58±3.86 343.12±112.49 17.37±2.44 434.13±942.77 20.00±3.53 89.88±89.88 Table 1. Survey results of Chimonobambusa szechuanensis and tree layer (Mean ± SD)
-
单因素方差分析的结果表明,乔木层郁闭度对八月竹发笋量的影响差异极显著(P<0.01),对一年生竹基径的影响差异显著(P<0.05);乔木株数对八月竹笋高度的影响差异极显著(P<0.01),对活竹株数、一年生竹高度和笋基径的影响差异显著(P<0.05);乔木平均高度对八月竹发笋量、活竹株数、死竹株数和一年生竹基径的影响差异极显著(P<0.01),对多年生竹基径的影响差异显著(P<0.05)(见表2)。
乔木层指标 八月竹指标 df F P 乔木层郁闭度 发笋量 df 1 =2, df 2 =234 6.926 0.002** 活竹株数 df 1 =2, df 2 =234 2.493 0.085 死竹株数 df 1 =2, df 2 =234 0.071 0.932 一年生竹高度 df 1 =2, df 2 =204 2.967 0.053 一年生竹基径 df 1 =2, df 2 =204 3.426 0.034* 多年生竹高度 df 1 =2, df 2 =234 0.759 0.469 多年生竹基径 df 1 =2, df 2 =234 0.759 0.469 笋高度 df 1 =2, df 2 =227 0.382 0.683 笋基径 df 1 =2, df 2 =227 0.382 0.683 乔木株数 发笋量 df 1 =2, df 2 =234 2.040 0.132 活竹株数 df 1 =2, df 2 =234 3.976 0.020* 死竹株数 df 1 =2, df 2 =234 1.084 0.340 一年生竹高度 df 1 =2, df 2 =204 3.804 0.024* 一年生竹基径 df 1 =2, df 2 =204 0.587 0.557 多年生竹高度 df 1 =2, df 2 =234 2.580 0.078 多年生竹基径 df 1 =2, df 2 =234 0.098 0.907 笋高度 df 1 =2, df 2 =227 7.053 0.002** 笋基径 df 1 =2, df 2 =227 3.582 0.029* 乔木平均高度 发笋量 df 1 =2, df 2 =234 44.710 <0.001*** 活竹株数 df 1 =2, df 2 =234 10.030 <0.001*** 死竹株数 df 1 =2, df 2 =234 4.808 0.009** 一年生竹高度 df 1 =2, df 2 =234 1.696 0.186 一年生竹基径 df 1 =2, df 2 =234 8.749 <0.001*** 多年生竹高度 df 1 =2, df 2 =234 0.341 0.711 多年生竹基径 df 1 =2, df 2 =234 3.350 0.037* 笋高度 df 1 =2, df 2 =227 1.220 0.297 笋基径 df 1 =2, df 2 =227 2.172 0.116 注:*为P<0.05,**为P<0.01,***为P<0.001。 Table 2. Variance analysis of growth index of Chimonobambusa szechuanensis in different groups of tree layer structure characteristics
-
根据方差分析结果,挑选对八月竹生长指标有显著性影响的因子,将其作为模型的固定效应。因此,可将乔木层郁闭度和乔木平均高度2个因子,保留在八月竹发笋量模型中;将乔木平均高度和乔木株数2个因子,保留在活竹株数模型中;将乔木平均高度因子保留在八月竹死竹株数模型中;将乔木株数因子保留在一年生八月竹平均高度模型中;将乔木层郁闭度和乔木平均高度2个因子,保留在一年生八月竹平均基径模型中;将乔木平均高度因子保留在多年生八月竹平均基径模型中;将乔木株数因子保留在八月竹笋高度和笋基径模型中。
采用赤池信息准则(AIC)指标来比较不同模型间的模拟效果。AIC值越小,表明模拟效果越好。结果表明,所挑选的主要因子模拟效果均好于没有显著性影响的因子的模型拟合效果。使用不同的有显著性的因子作为固定效应,其混合模型的精度不同。在八月竹发笋量模型中,以乔木平均高度作为固定效应的模型AIC值最小,拟合精度最高;在八月竹发笋量、活竹株数、死竹株数、一年生平均基径、多年生平均基径模型中,以乔木平均高度作为固定效应的模型AIC值均最小,拟合精度均最高;在一年生平均高度、笋基径和笋高度模型中,以乔木株数作为固定效应的模型AIC值均最小,拟合精度均最高(见表3)。
八月竹指标 乔木层指标 固定效应截距 固定效应系数 随机效应截距 残差 AIC 发笋量 乔木层郁闭度 1.7619*** −0.1228 0.0929 — 455.4 乔木平均高度 2.1990*** −0.0441* 0.0657 — 450.8 活竹株数 乔木平均高度 2.2924*** −0.0067 0.2207 — 1347.3 乔木株数 2.2376*** −0.0008 0.2211 — 1347.4 死竹株数 乔木平均高度 2.9551*** 0.0026 0.0577 — 1569.7 一年生平均高度 乔木株数 316.4935*** 1.1803 9290.9000 2838.4000 2379.0 一年生平均基径 乔木层郁闭度 15.9130*** 0.4150 12.3466 3.5372 1015.8 乔木平均高度 15.1459*** 0.0995 12.3776 3.5370 1015.1 多年生平均基径 乔木平均高度 17.5449*** 0.0259 6.4550 1.9490 991.1 笋高度 乔木株数 136.0558*** 0.3232 4943.5000 3816.6000 2621.3 笋基径 乔木株数 19.0495*** 0.04883** 6.3984 5.8990 1161.6 注:*为P<0.05,**为P<0.01,***为P<0.001。 Table 3. Model fitting results of Chimonobambusa szechuanensis
结果显示,八月竹发笋量、活竹株数、死竹株数、一年生平均高度、一年生平均基径、多年生平均基径、笋高度和笋基径模型的固定效应截距均达到极显著水平(P<0.001),说明乔木层结构特征各参数对模型变化影响极显著。乔木平均高度的固定效应系数对八月竹死笋株数模型变化影响极显著(P<0.01),乔木株数的固定效应系数对八月竹笋基径模型变化影响极显著(P<0.01),乔木平均高度的固定效应系数对八月竹发笋量模型变化影响显著(P<0.05)(见表3)。
随着乔木层郁闭度和乔木平均高度的增加,八月竹发笋量均呈减少的趋势;随着乔木平均高度和乔木株数的增加,活竹株数均呈减少的趋势;随着乔木平均高度的增加,八月竹死竹株数呈增加的趋势;随着乔木株数的增加,一年生八月竹平均高度呈增加的趋势;随着乔木层郁闭度和乔木平均高度的增加,一年生八月竹平均基径均呈增加的趋势;随着乔木平均高度的增加,多年生八月竹平均基径呈增加的趋势;随着乔木株数的增加,八月竹笋高度和笋基径均呈增加的趋势(见表3)。
-
竹子和木本植物是大熊猫栖息地的重要组成部分[17],竹子生长状况与木本植物之间具有关系[18]。大熊猫、森林和竹子三者协同进化[19]。在大熊猫国家公园荥经片区羊叉岗旧苗圃基地和码棚头原生森林生态系统林下分布有八月竹的区域,利用样线法和样方法,开展森林群落乔木层与林下八月竹调查,并利用GLMM和LMM模型,构建了大熊猫栖息地森林群落乔木层结构特征与林下八月竹生长指标的关系,这为大熊猫栖息地恢复提供了定量的数据支撑。
修复效果、修复要求和修复考虑因素是大熊猫栖息地修复的主要研究主题[20]。竹子的空间分布对于大熊猫栖息地的分布具有重要影响[21],竹子密度、基径、高度和幼竹所占比例是衡量森林中竹子资源的重要指标[22],也是表征竹子生长状况的指标[18]。大熊猫的栖息地选择对获得高可用性的美味竹子和良好的庇护所具有重要的影响[23]。幼竹的可利用性是竹林生境选择的重要驱动因素[24],竹林密度、高度、基径、竹笋的生长、出笋量、无性系根茎等,均受温度、湿度、光照强度等环境因子的显著影响[25]。乔木作为森林群落的最高层,其郁闭度的大小会对林下环境(如光照强度、温度、湿度等)产生影响,从而影响竹子的生长和天然更新。同一群落内缺苞箭竹(Fargesia denudata)天然更新受冠层郁闭度和层片结构影响[26];不同乔木郁闭度下竹子生长和更新往往存在差异[27]。本研究结果表明,八月竹发笋量随乔木层郁闭度增加而减少,这与黄荣澄等对八月竹的研究结果[28]以及魏辅文和冯祚建对峨热竹(Arundinaria spanostachya)的研究结果[29]一致。随着乔木平均高度增加,乔木树冠逐渐变大且浓密,乔木层郁闭度也随之增大。因此,八月竹发笋量也随乔木平均高度增加而减少,此研究结果与Hong对峨热竹的研究结果[30-32]一致,同时也暗示了高郁闭度或者较弱的光照可以减少八月竹发笋量。不同郁闭度对不同竹种的生长更新的影响不同,如全光照下细枝箭竹(Fargesia stenoclada)有较高的密度、基径和高度[27]。本研究中,活竹株数与乔木株数和乔木平均高度均呈负相关关系,而死竹株数与乔木平均高度呈正相关关系,这可以解释为随着乔木株数和乔木平均高度增加,乔木层郁闭度增加,过分荫蔽的环境影响根系的发育,使竹子幼苗的根系生长脆弱,根系竞争能力差,影响幼苗对水分和养分的吸收,造成幼苗死亡[33]。已有研究表明,缺苞箭竹的分株生物量、基径、高度以及分株各构件生物量随林冠郁闭度减小均表现为先增加后减小的趋势,在小林窗中达到最大值[34];华西箭竹(Fargesia nitida)的平均高度随着乔木层郁闭度的增加而减少[35,36]。而本研究中,一年生八月竹平均高度与乔木株数呈正相关关系,一年生八月竹平均基径与乔木层郁闭度和乔木平均高度均呈正相关关系,多年生八月竹平均基径与乔木平均高度也呈正相关关系,这可能是因为竹种不同,竹子对乔木层的适应也有所不同,也可以解释为乔木层郁闭度随着乔木株数和乔木平均高度的增加而增加,郁闭度较高会促进八月竹个体基径和高度的生长,也体现了不同克隆植物对应环境异质性策略的差异。八月竹笋高度和基径均随着乔木株数增加而增加,这也说明了郁闭度较高可以促进八月竹笋的生长。
大熊猫喜欢活动于上层有乔木、下层有竹林的环境中,不同的山系大熊猫喜欢的乔木层郁闭度不同,但基本不利用竹子密度过高的纯竹林[37]。大熊猫国家公园荥经片区是大相岭山系大熊猫种群的核心分布和集中分布区,也是连接大相岭山系和邛崃山山系大熊猫种群基因交流的关键区。区内有大片上层无乔木层覆盖的八月竹纯林,不属于大熊猫理想的栖息地。对大熊猫国家公园荥经片区大熊猫栖息地进行恢复是恢复大相岭山系大熊猫栖息地的重要基础,是保护、维持大熊猫种群数量的有效途径。因此,要为该区域大熊猫创造良好的栖息地环境,可以将森林和竹子之间的关系,作为大熊猫栖息地人工恢复的参考标准。通过研究了解了大相岭山系大熊猫栖息地森林群落乔木层结构特征与林下八月竹的生长指标之间的关系,可以科学合理的采取经营措施,如通过栽种本土阔叶树种,提高乔木层郁闭度,从而控制林下八月竹无性系种群数量,降低八月竹林密度,以确保合理的乔木层结构特征促进大熊猫取食竹生长更新。
Study on the Relationship between Tree Layer and Understory Chimonobambusa szechuanensis in Giant Panda Habitat
doi: 10.12172/202212220001
- Received Date: 2022-12-22
- Available Online: 2023-10-10
- Publish Date: 2023-12-25
-
Key words:
- Giant panda habitat /
- Habitat restoration /
- Forest canopy /
- Chimonobambusa szechuanensis /
- Generalized linear mixed model
Abstract: In order to explore the correlation between forest community and understory giant panda feeding bamboo in the giant panda habitat of the Daxiangling mountain, the tree layer structure characteristics of forest community and the growth index of Chimonobambusa szechuanensis were investigated, and the generalized linear mixed model (GLMM) and linear hybrid model (LMM) were constructed and analyzed. The results showed that: (1) The canopy density of tree layer had significant effects on the number of bamboo shoots and the diameter of annual bamboo base; the number of trees had significant effects on the number of Chimonobambusa szechuanensis, height of annual bamboo, height of bamboo shoots and base diameter of bamboo shoots; the average height of trees had significant effects on the number of shoots, number of Chimonobambusa szechuanensis, number of dead bamboos, annual and perennial bamboo base diameter. (2) the number of bamboo shoots decreased with the increase of canopy density and average tree height; the number of living bamboos decreased with the increase of the number and average height of trees; the number of dead bamboos increased with the increase of average height of tree; the average height of bamboo, the height of bamboo shoots and the base diameter of bamboo shoots increased with the number of trees; the average base diameter of annual and perennial August bamboo increased with the increase of average tree height.
























 DownLoad:
DownLoad: