






 下载:
下载:




-
蒜头果(Malania oleifera)为铁青树科蒜头果属植物,是我国特有的单种属植物,属国家二级濒危保护物种。蒜头果分布范围小,仅分布于中国云南省广南县、富宁县和广西壮族自治区的龙州、靖西、德保、大新等县市的狭窄地带[1]。蒜头果种仁油含量高达65%,油中含有高达45%左右的神经酸(Nervonic acid,NA)[2]。神经酸是一种超长链单不饱和脂肪酸(C24:1,cis-15-tetracosenic acid),是大脑神经纤维细胞膜中鞘磷脂的核心天然成分,具有预防和治疗老年痴呆症等神经退行性疾病的功效[3]。蒜头果生存的自然环境特殊,主要生长在喀斯特石灰岩山地,对干旱的石漠化生境的生态适应性强,是石漠化治理的重要树种。蒜头果分布区海拔落差大,从500米到1400米均有分布。立地条件差异显著,以岩石裸露率70—90%的石山和岩石裸露率小于20%的土山为主,石山为土层薄, 土壤干燥的石灰岩山地,土山为土壤层厚度达30 cm 的沙岩和红壤山地[4-5]。研究表明,不同立地条件蒜头果种子的油含量和神经酸含量存在一定差异,石山生长的蒜头果种子油含量和神经酸含量均高于土山[6-7],其机理尚不清楚。
植物的生长和生理活动受到多种营养元素的调节,植物生长的必需营养元素有16种,包括N、P、K等大量元素,Ca、Mg、S等中量元素和Zn、Fe、Mn等微量元素[8]。元素通过单一含量或者不同元素之间的平衡调节植物的不同发育过程和生理活动。土壤是植物生长发育的物质基础,也是植物营养的主要来源,植物的根系从土壤中获得大量的养分,利于树木体内各种物质的合成和各种营养物质的积累。营养元素在土壤中以不同形式存在,有些元素的形式不经转化无法被植物吸收利用。土壤微生物的分解作用,能将植物的残根、烂根、落叶等分解转化为营养元素供给于植物。同时微生物的代谢产物还可以分解和转化矿物质,将植物和土壤中的矿质元素提供给植物吸收利用,微生物的生命活动在中量和微量矿质营养元素的转化中起着十分重要的作用[9]。
对分布于云南省广南县和富宁县不同居群成年蒜头果植株的土壤理化性质以及土壤微生物进行了检测分析,并对石山和土山的元素和微生物特征进行了比较分析,旨在探究与蒜头果种子油含量及神经酸含量相关的营养元素和土壤微生物类型。
-
云南省广南县和富宁县是蒜头果的主要自然分布区,广南县(104º31′—105º39′ E,23º29′—24º28′ N)位于云南省东南部,境内蒜头果分布区为岩石裸露率大于70%的石山生境,土壤以石灰岩土为主[10-11],富宁县(105º14′—106º13′ E,23º31′—24º09′ N)位于云南省东南部,境内蒜头果分布区为岩石裸露率约5%的土山生境,其蒜头果分布区土壤以红壤为主[12]。研究对象为广南县集中分布的三个居群和富宁县集中分布的两个居群,每个居群采集三个成年植株的土壤进行分析(见表1)。
表 1 样地概况
Table 1. Basic status of plots
地点 经度 纬度 海拔/m 生境 (岩石裸露率) 样本数 平均胸径(cm) 平均树高(m) 旧莫乡(JM) 104°90'E 23°92'N 1278.25 石山 (90%) 3 30.6 7.5 八宝镇(BB) 105°43'E 23°86'N 1027.49 石山 (70%) 3 21.8 8.5 曙光乡(SG) 105°16'E 23°72'N 1366.18 石山 (80%) 3 27.8 8.7 者桑乡(ZS) 106°08'E 23°74'N 444.04 土山 (5%) 3 20.7 12.8 板仑乡(BL) 105°73'E 23°65'N 843.00 土山 (5%) 3 24.9 9.5 -
2021年9月,在云南文山州广南县的旧莫乡(JM)、八宝镇(BB)、曙光乡(SG)和富宁县的者桑乡(ZS)、板仑乡(BL)等5个蒜头果的分布点,共选取生长健壮的野生成年植株15株以供研究,其中石山生境居群三个共9株,土山生境居群2个共6株。土壤采样主要采集根际的表土层(0—15 cm),去除土壤表层的凋落物后,以蒜头果植株为中心,在离植株基部25 cm的位置延东、南、西、北四个方位,紧沿着蒜头果的根部采集表层土,再将四个位置的土样混合均匀,并去除石砾、动植物残体及杂质后过2 mm筛,共计15个土壤样品,分成两份分别装入自封袋并标记,一份自然风干后用作土壤理化性质测定,一份保存在−20℃冰盒用于土壤微生物磷脂脂肪酸测定。
-
土壤pH值使用pH计(FE-20,梅特勒-托利多仪器(上海)有限公司)测定;土壤有效磷(AP)、硝态氮(NO3−-N)、铵态氮(NH4+-N)用盐酸氟化铵浸提后,滤液用连续流动分析仪(Auto Analyzer 3,SEAL Analytical GmbH)测定;土壤全碳(C)、全氮(.N)采用元素分析仪分析仪(Vario MAX CN,Elementar Analys ensysteme GmbH)测定。土壤全磷(P)、全钾(K)、全钙(Ca)、全镁(Mg)、全硫(S)、全铁(Fe)、全锰(Mn)、全铜(Cu)、全锌(Zn)、全硼(B)、全钠(Na)、全铝(Al)、全硅(Si)用HNO3-HClO4消解后过滤,滤液用电感耦合等离子体发射光谱仪(iCAP6300,Thermo Fisher Scientific U.S.A)测定。
土壤微生物磷脂脂肪酸测定使用气相色谱仪(Hewlett-Packard 6890 series GC, FID)上采用MIDI软件系统(MIDI, Inc., Newark, DE)进行分析,测定磷脂脂肪酸(PLFA)各组分的含量。以所报道的磷脂脂肪酸标记(Table S1)为参照统计分析土壤革兰氏阳性菌(GP)、革兰氏阴性菌(GN)、放线菌(ACT)、丛枝菌根真菌(AMF)、腐生真菌(SF)、真菌总量(Fungi)、细菌总量(Bacteria)。F: B代表Fungi: Bacteria,A: B代表AMF: Bacteria[13-14]。
-
用 Microsoft Excel 2010 软件进行数据的初步整理和对数转换后,采用Origin Pro(10.0)软件对不同居群土壤理化性质和土壤微生物组成进行方差分析,运用SPSS(25.0)软件中的独立样本t检验分析不同生境的土壤理化性质和土壤微生物;采用R(4.3.1)软件的主成分分析(PCA)探究土壤理化性质和土壤微生物的差异;运用Spearman相关分析法探究土壤理化性质和土壤微生物群落之间的相关关系,利用Origin Pro(10.0)作图。
-
通过对蒜头果5个居群(JM、BB、SG、ZS、BL)的土壤理化性质和土壤微生物特征的分析。结果表明,蒜头果植株不同居群间土壤pH值有显著差异,JM(7.4)、SG(5.6)的土壤pH值显著高于BB(4.6)、ZS(4.5)BL(4.4),JM的土壤pH值显著高于SG(P < 0.05)。大量元素C、N、P、K的平均含量分别为49.898、4.145、1.092、12.713 g·kg−1,其中K元素含量较高,且在居群间差异较大;中量元素Ca、Mg、S的平均含量分别为3.927、4.413、0.301 g·kg−1,JM居群Ca元素含量显著高于其他居群,Mg元素在居群间差异较大,JM、BB、SG居群显著高于ZS、BL居群;微量元素Fe、Mn、Cu、Zn、B的平均含量分别为33.209、1.546、0.031、1.092、0.119、0.057 g·kg−1,其中Fe元素含量最高,且表现为JM、BB、SG居群显著高于ZS、BL居群(P < 0.05);NH4+-N、NO3−-N、AP含量在不同居群间没有显著差异(P > 0.05)(见图1A)。
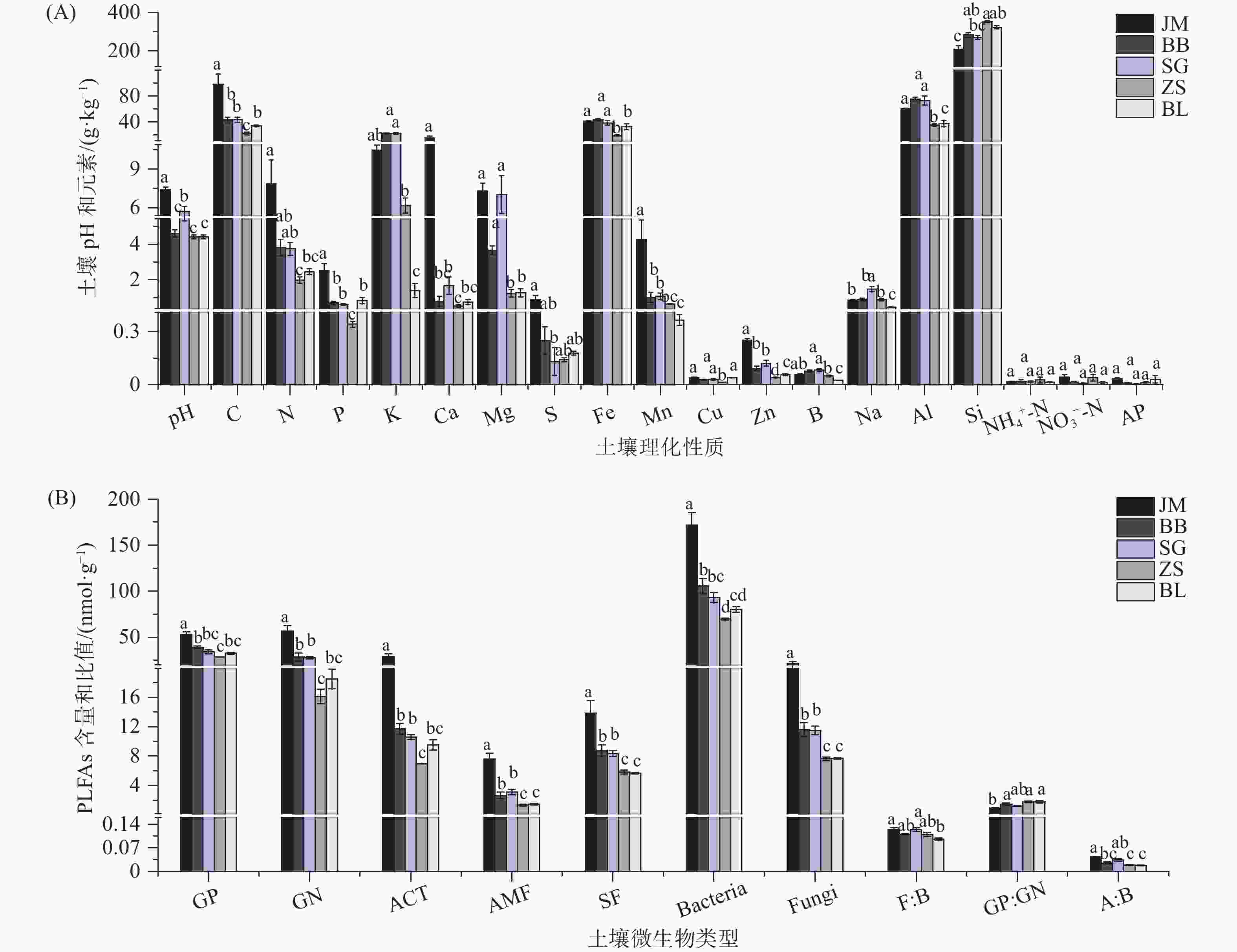
图 1 不同居群的蒜头果土壤理化性质(A)以及土壤微生物生物量(B)的差异(均值 ± SE)。不同小写字母表示蒜头果居群间的差异(P < 0.05)。
Figure 1. Physical and chemical properties of the soil (A) and soil microbial biomass (B) of Malania oleifera in different populations (Mean ± SE). Different lowercase letters indicate significant Malania oleifera difference populations (P < 0.05).
已有研究表明,蒜头果在不同的立地条件下,油含量和神经酸有明显的差异(赖家业, 1999),石山居群的蒜头果油含量和神经酸含量均高于土山居群[7]。各种土壤理化特性中,土壤养分对植物的生长影响最大[16]。研究中K、Mg、Fe等3种元素和土壤水分具有较为一致的空间分布规律,含量高值区主要集中于研究区广南县;这可能是因为JM、BB、SG居群的土壤类型均为砾石质,能够减少水分蒸发,提高土壤保水能力,有利于土壤养分转化与循环,从而有效的被植物吸收,而且JM、BB、SG具有高的土壤养分和水分,对种群的更新和植物生长非常有利。因此这表明K、Mg、Fe元素和土壤水分是影响蒜头果油含量和神经酸含量的潜在因子。
蒜头果植株不同居群土壤微生物的平均生物量为GP(37.273 nmol·g−1)、GN(29.450 nmol·g−1)、ACT(13.494 nmol·g−1)、AMF(3.234 nmol·g−1)、SF(8.499 nmol·g−1)、Bacteria(104.036 nmol·g−1)、Fungi(11.973 nmol·g−1),F: B、GP: GN、A: B的平均比值为0.113、1.433、0.028(图1B),特别的是JM、BB、SG的AMF、SF、Fungi生物量显著高于ZS、BL(P < 0.05)。土壤微生物是土壤的重要组成部分,在推动土壤养分转化、物质循环和能量流动过程中发挥了重要作用,其结构和功能对周围环境变化特别敏感,常作为土壤养分状况的重要指示[17-18]。研究发现,真菌类的土壤微生物(AMF、SF、Fungi)在JM、BB、SG居群表现出高生物量。AMF是一种存在于根际土壤中,能与陆地上大多数维管植物形成互利互惠的关系的菌根真菌,研究表明其独特的丛枝结构,能增加植物对水分和矿质元素的吸收,土壤 pH、湿度和透气性能直接影响发育早期菌根真菌的生长繁殖[19]。JM、BB、SG居群的AMF生物量显著高于ZS、BL,而JM、BB、SG居群的土壤疏松、肥沃,适合 AMF生长,故AMF 的生物量高于ZS、BL,这说明AMF的生物量受到立地条件的影响。植物组织的物理分解先由蚯蚓和螨虫等分解,然后腐生真菌再分解纤维素、木素和其他复杂大分子,最后所得化合物将被细菌进一步处理,因此,腐生真菌通常被认为是分解过程的主要引擎[20]。腐生真菌作为土壤碳循环的重媒介,受土壤理化性质的调控[21]。研究表明土壤全氮的积累会直接影响腐生真菌类群的聚集[22]。在我们的研究中,JM、BB、SG居群的土壤N含量高于ZS、BL,可能是JM、BB、SG的土壤是砾石质,养分高,这对土壤全氮的积累有积极影响,是JM、BB、SG居群腐生真菌类群聚集的原因之一。土壤的pH和养分含量会影响真菌的生长和繁殖,直接改变土壤中真菌群落组成[23],研究表明真菌生物量与pH和养分含量成正比[24]。研究中不同居群的土壤pH值、元素含量与真菌生物量具有一致的空间规律,说明蒜头果油含量和神经酸含量存在差异的主要微生物是真菌。
-
不同居群的蒜头果土壤微生物与土壤理化性质之间关系紧密(见图2)。AMF、SF、Fungi与pH、K、Mg、Fe呈显著正相关(P < 0.05)。研究表明,土壤微生物的生物量与土壤理化性质之间存在关联,微生物所需的养分主要来源于土壤中的养分。当土壤营养物质丰富时,微生物群落的数量、生物量等与土壤N等呈正相关,说明丰富的营养物质大大促进了微生物的定居、生长和繁殖[25]。研究中,土壤各微生物生物量与土壤pH、K、Mg、Fe呈极显著正相关,这与大多数研究结果一致。此外,我们发现相对于ACT,SF和B,pH对AMF和F的影响更加显著,而对GP的影响最弱。所以土壤pH、K、Mg、Fe、AMF、F可能是导致不同居群蒜头果油含量和神经酸含量差异的主要因子。
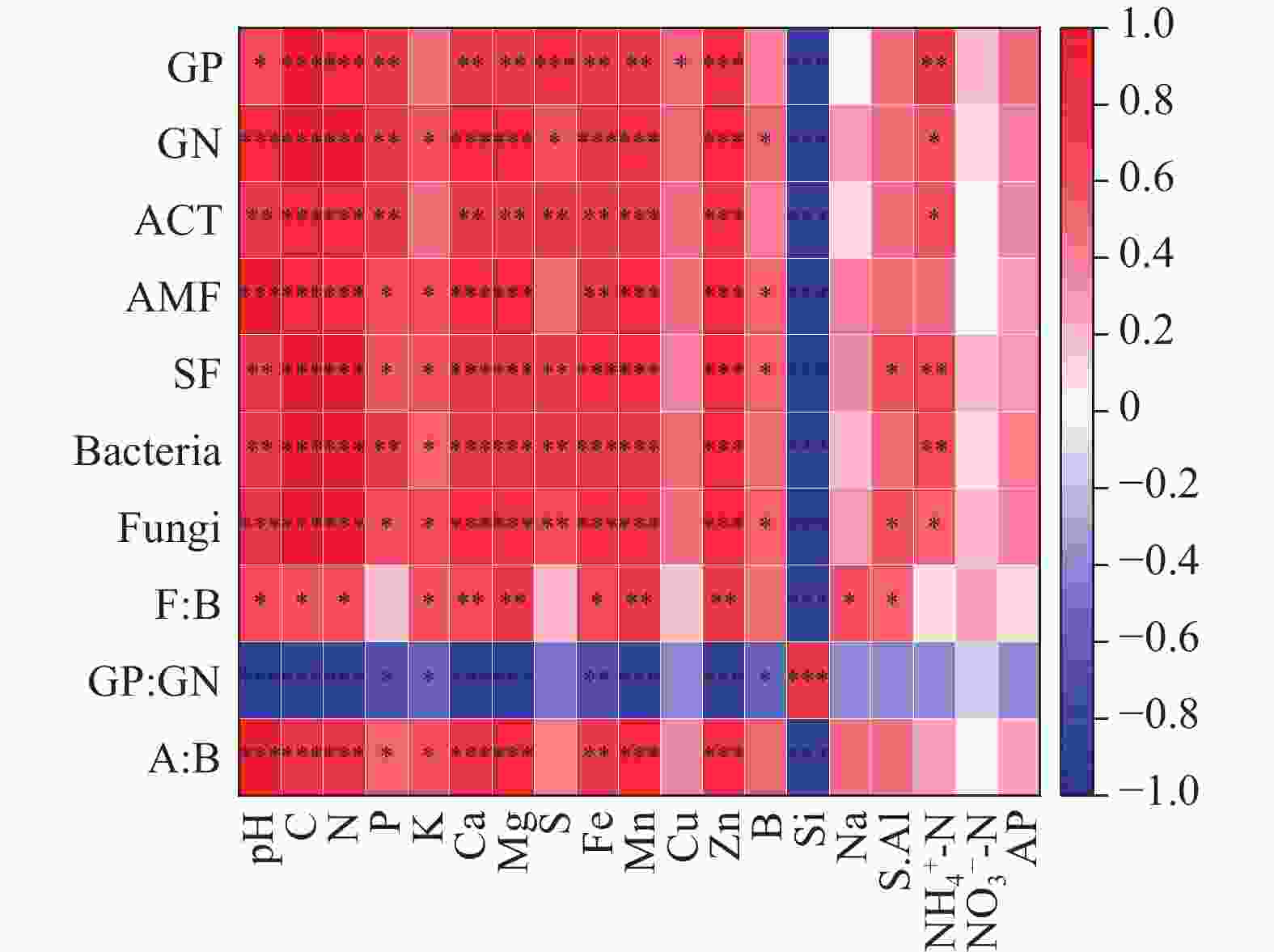
图 2 不同居群间蒜头果土壤理化性质和土壤微生物生物量的 Spearman 相关性分析。注:*P < 0.05;**P < 0.01;***P < 0.001。
Figure 2. Results on bivariate relationships on physical and chemical properties and soil microbial biomass of Malania oleifera in idifferent populations using Spearman correlation. Note: *P < 0.05; **P < 0.01; ***P < 0.001.
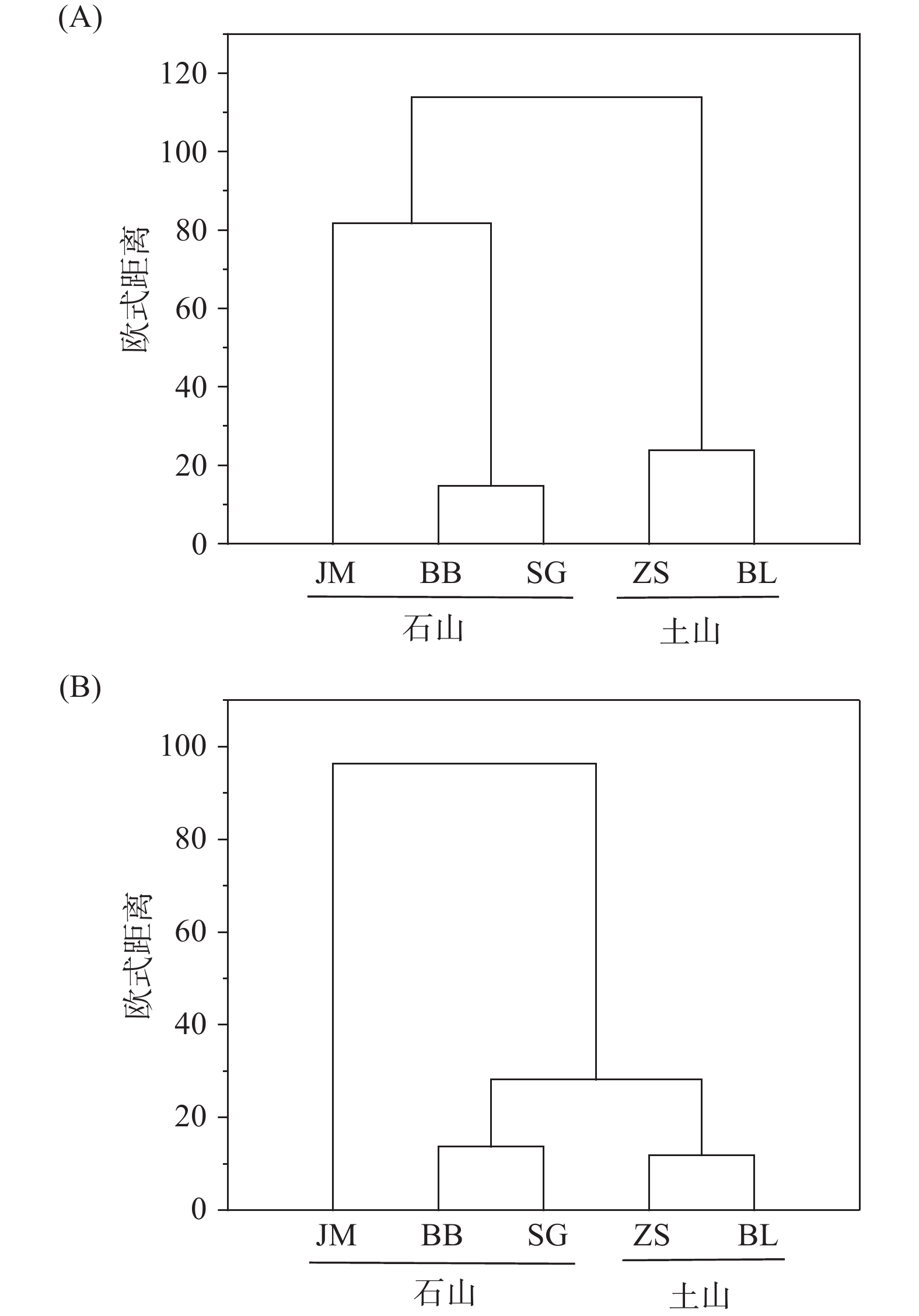
以蒜头果不同居群的土壤理化性质值和土壤中PLFA 的生物量为指标构建矩阵,以欧式距离为尺度,用最小距离法进行系统聚类分析。结果显示,5个蒜头果居群的土壤理化性质主要分为2大类群(见图3A),JM、BB、SG聚为第一类群;第二类群为ZS、BL;其中,第一类群又可进一步细分为2个亚类群,分别为JM和BB、SG;显而易见的是,BB、SG这个两个居群的遗传亲缘性较高。5个蒜头果居群的土壤微生物主要分为3大类群(见图3B),JM聚为第一类群;第二类群为BB、SG;第三类群为ZS、BL;其中BB、SG、ZS、BL这个四个居群的遗传亲缘性较高。
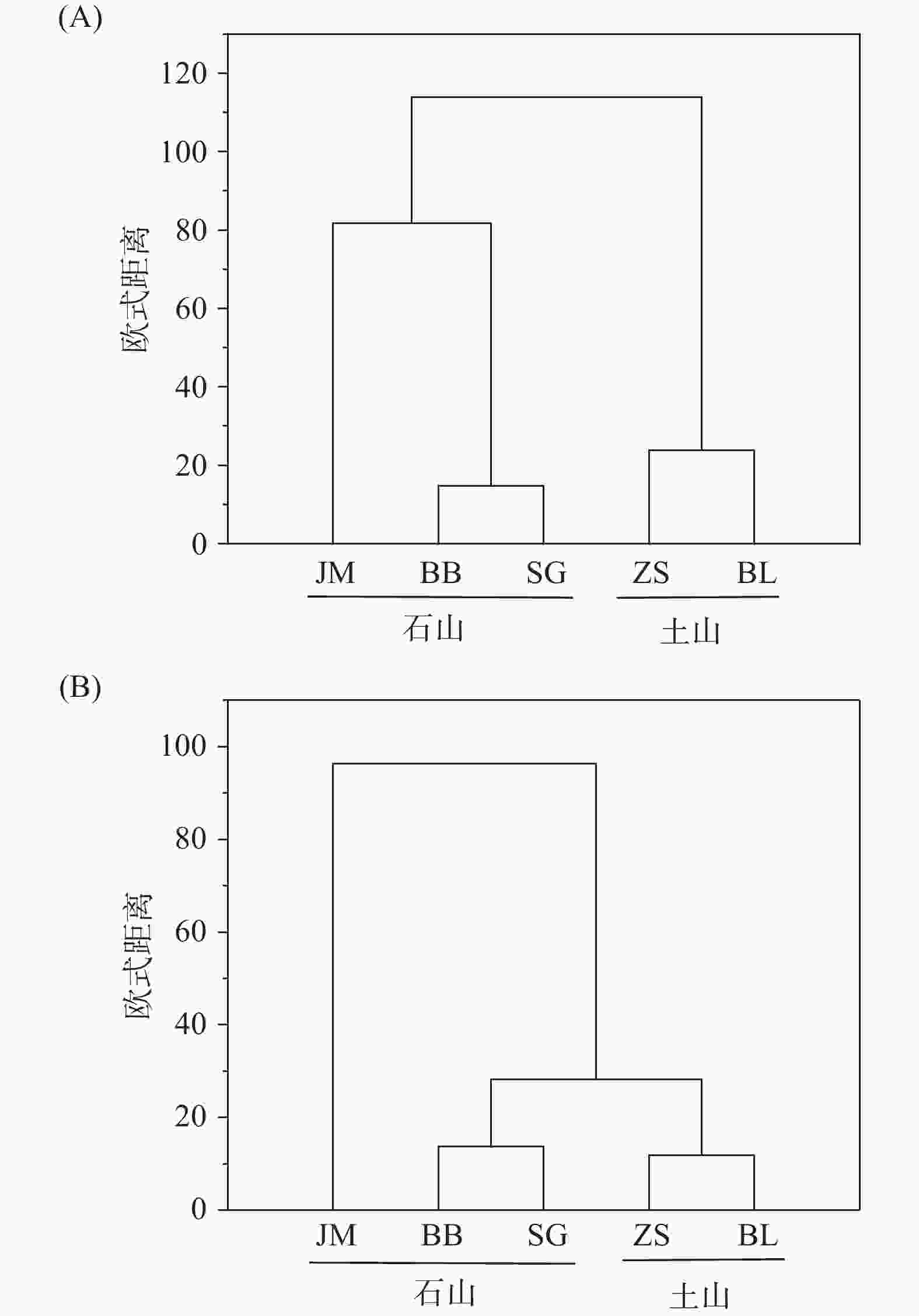
图 3 不同居群蒜头果土壤理化性质(A)和土壤微生物生物量(B)的聚类分析。
Figure 3. Cluster analysis on physical and chemical properties (A) and soil microbial biomass (B) of Malania oleifera in idifferent populations.
聚类分析结果表明,JM、BB、SG的元素特征较为相似,这三个居群为岩石裸率较高的石山生境,ZS和BL的元素特征较为相似,这两个居群为岩石裸露率较低的土山生境。土山居群与石山居群的元素特征存在较大差异;而土壤微生物组成同样表现为石山居群与土同居群存在一定的差异,但JM居群岩石裸露率高达90%,微生物组成与另两个石山居群BB和SG存在一定差异,表明土壤微生物组成受立地条件影响较大。JM、BB、SG的土壤pH值、元素含量和土壤微生物生物量比ZS、BL高,这可能也是不同居群的蒜头果油含量、神经酸含量存在差异的原因之一。
-
将不同居群按石山生境与土山生境两个不同的立地条件进行比较分析,两个立地条件下的蒜头果植株土壤pH值有显著差异,石山生境的土壤pH值(6.0)显著高于土山生境(4.4)(P < 0.05)。
大量元素C、N、P、K的平均含量分别为石山(64.401、5.387、1.316、18.328 g·kg−1)、土山(28.139、2.282、0.757、4.290 g·kg−1),其中K元素含量较高,且在生境间差异较大;中量元素Ca、Mg、S的平均含量分别为石山(6.052、3.380、0.401 g·kg−1)、土山(0.739、1.463、0.151 g·kg−1),石山生境的Ca元素含量显著高于土山生境,Mg元素也在生境间差异较大;微量元素Fe、Mn、Cu、Zn、B的平均含量分别为石山(41.140、2.243、0.034、0.164、0.069 g·kg−1)、土山(21.312、0.501、0.027、0.052、0.038 g·kg−1),其中Fe元素含量最高,且表现为石山生境显著高于土山生境(P < 0.05);而石山的Si元素含量显著低于土山(P < 0.05),NH4+-N、NO3−-N、AP含量在不同居群间没有显著差异(P > 0.05)(图1 A)。
这表明石山生境的土壤肥力较高,与前人研究的不同立地条件的蒜头果土壤pH和养分差异的结果一致[15]。石山的土壤因母岩性质多表现为富含K、Ca、Mg等特性,生长在石灰岩的植物多表现出高Ca、高Mg的特征,一些专性钙生植物对这类土壤表现出高度依赖,这也从侧面表现出石灰岩土壤矿质元素对植物生长发育的限制性[26]。研究表明高度风化的土壤具有较低的Si含量[27-28],这与研究结果一致。石山的K、Mg元素含量明显高于滇南喀斯特地区,Fe元素含量明显低于滇南喀斯特地区,这可能与蒜头果分布有关[29]。由此推测,蒜头果种子的油含量和神经酸含量差异与生境中土壤pH和K、Ca、Mg、Fe、Si等元素导致的。
蒜头果植株石山和土山生境的土壤微生物的平均生物量为GP(41.835 、30.430 nmol·g−1)、GN(37.560、17.284 nmol·g−1)、ACT(16.995、8.244 nmol·g−1)、AMF(4.459、1.396 nmol·g−1)、SF(10.324、5.761 nmol·g−1)、Bacteria(123.481、74.870 nmol·g−1)、Fungi(14.842、7.669 nmol·g−1),平均比值为F: B(0.119、0.103)、GP: GN(1.202、1.781)、A: B(0.034、0.019)(Fig.1B),在不同生境,石山的AMF、SF、Fungi生物量同样显著高于土山(P < 0.05)。
这可能是石山生境中的植物根系及凋落物分布比土山生境多,改变了土壤的碳输入,使石山的土壤微生物获得的碳源更加充沛,所以石山的土壤微生物种类和含量较丰富。研究表明土壤营养丰富意味着微生物含量亦丰富[30],研究中石山的多种元素含量显著高于土山境,所以石山具有较高的土壤微生物含量。在土壤中植物根系分泌物为微生物提供营养和能量,微生物大量聚集根系周围通过分解有机营养(促进养分的循环)及分解为植物提供养分,活化土壤中无效的矿质营养元素,并且通过合成植物激素影响植物的生长[31],所以土壤微生物可能会影响蒜头果种子的油含量和神经酸含量,具体是哪种微生物,需要进一步验证。
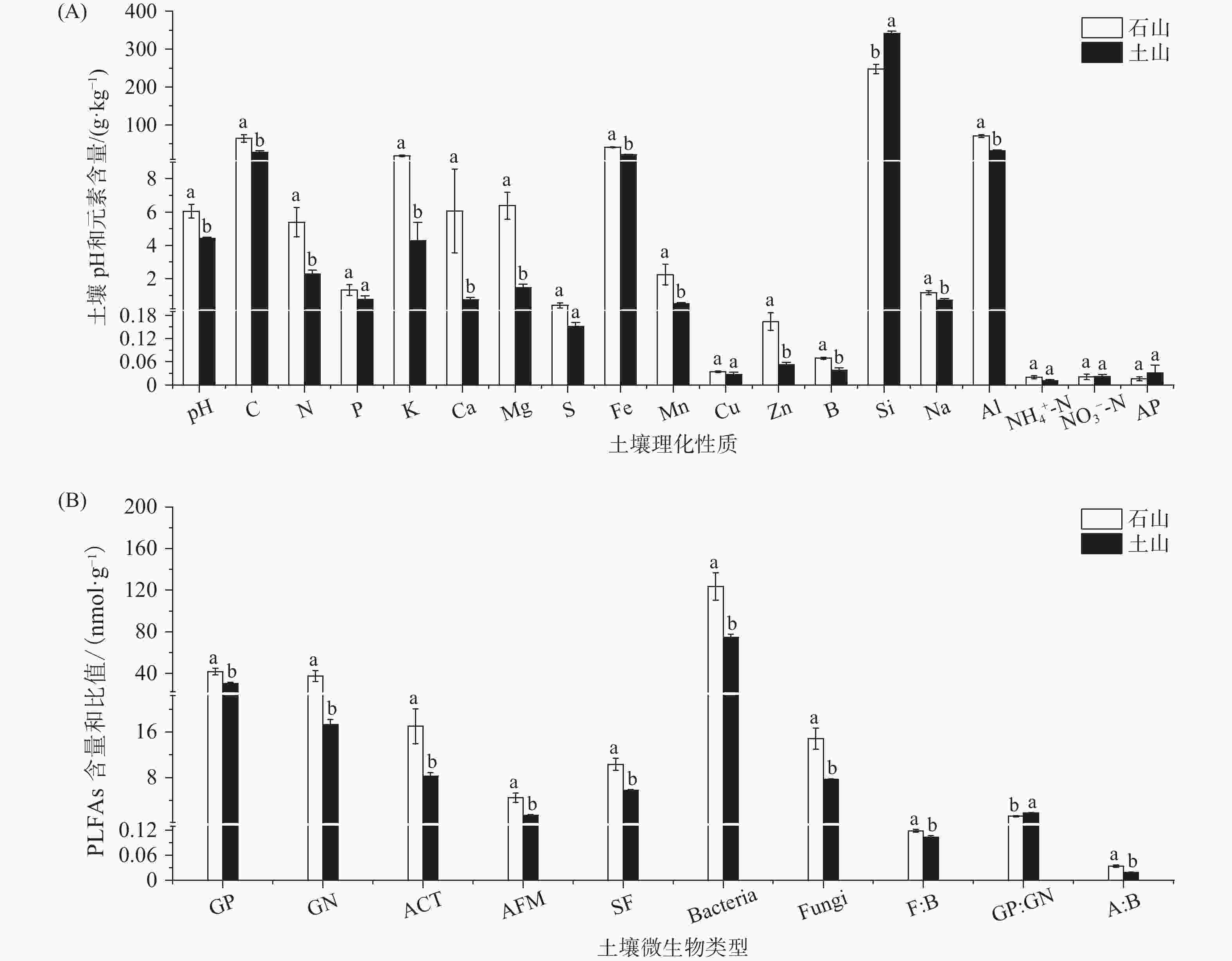
图 4 不同生境的蒜头果土壤理化性质(A)以及土壤微生物生物量(B)的差异(均值 ± SE)。不同小写字母表示蒜头果生境间的差异(P < 0.05)。
Figure 4. Physical and chemical properties of the soil (A) and soil microbial biomass (B) of Malania oleifera in different habitats (Mean ± SE). Different lowercase letters indicate significant Malania oleifera difference habitats (P < 0.05).
-
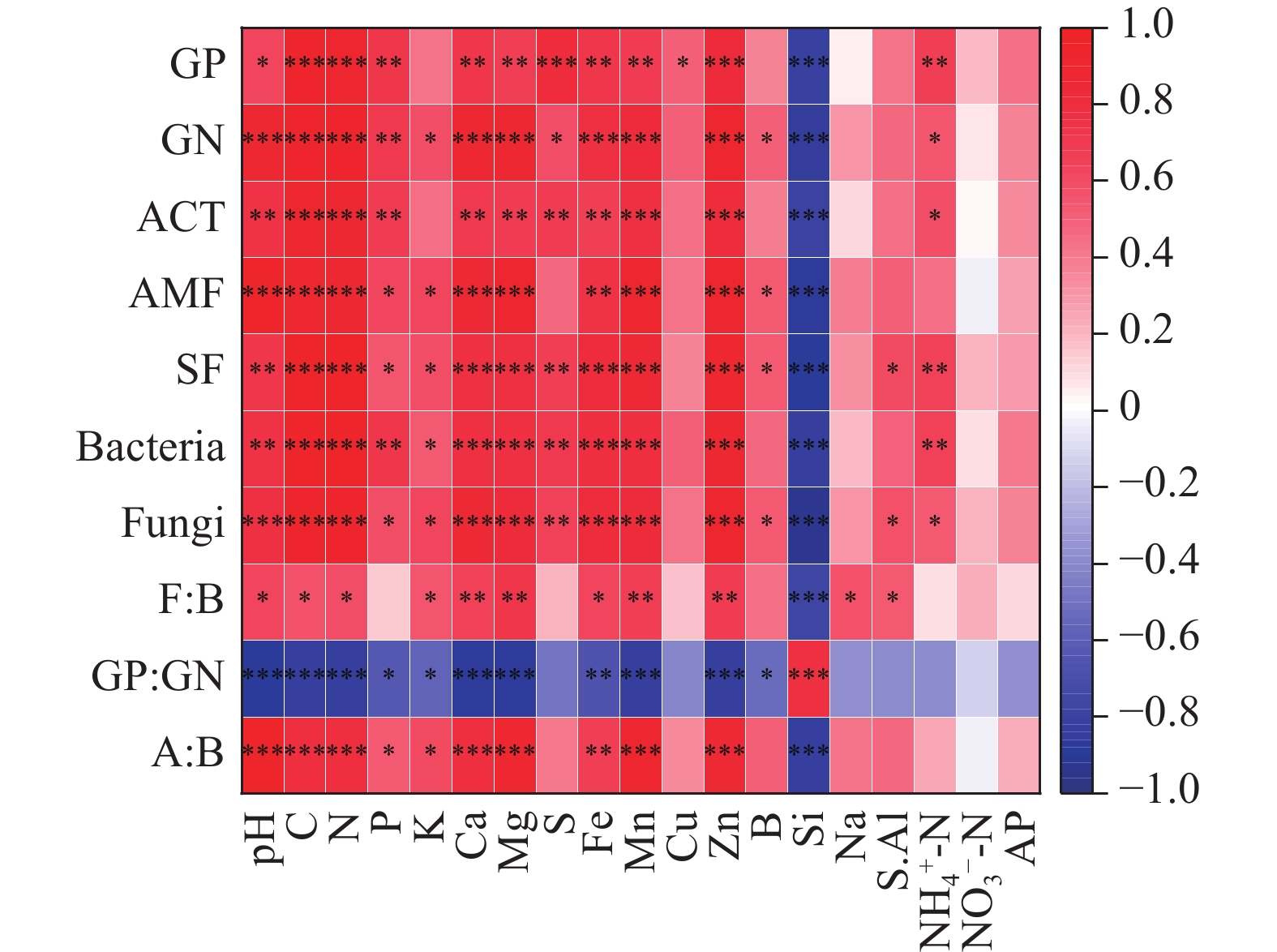
不同生境的蒜头果土壤理化性质与土壤微生物之间关系紧密(见图5)。AMF、SF、Fungi与pH、K、Mg、Fe呈显著正相关(P < 0.05)。土壤养分是微生物群落稳定的能量和营养来源,是土壤微生物群落形成的必要因素;另一方面,土壤微生物通过凋落物分解及与植物根系分泌物的相互作用,进一步促进土壤养分的积累,是土壤改良的主要动力[31]。研究中蒜头果各类土壤微生物与土壤C、N、Ca、Mg、Fe、Mn、Zn、Si元素极极显著相关,说明这些元素是影响微生物生长和发育的重要因素,而土壤Si与GP、GN、ACT、AMF、SF、Bacteria、Fungi呈极显著负相关关系,说明土壤Si是土壤微生物群落组成差异的主要驱动因子。

图 5 不同生境间蒜头果土壤理化性质和土壤微生物生物量的Spearman相关性分析。注:*P < 0.05;**P < 0.01;***P < 0.001。
Figure 5. Results on bivariate relationships on physical and chemical properties and soil microbial biomass of Malania oleifera in idifferent habitats using Spearman correlation. Note: *P < 0.05; **P < 0.01; ***P < 0.001.
通过主成分分析法(PCA)分别对蒜头果土壤理化性质及土壤微生物群落进行了综合分析(见图6)。从蒜头果不同生境的土壤理化性质来看(见图6A),第1主成分的方差贡献率为51.13%,第2主成分的方差贡献率为25.63%,石山和土山的土壤理化性质存在显著差异,主导因子是pH、Zn、K、Si,这可能是因为地理位置不同,小环境条件不同,导致蒜头果两类生境的土壤理化性质有显著差异。而对于蒜头果的土壤微生物群落(见图6B),第1主成分的方差贡献率为87.11%,第2主成分的方差贡献率为8.08%,主导因子是AMF、SF、Fungi,石山和土山的土壤微生物群落结构十分相似,这与张仲富[32]等人研究蒜头果根际微生物的结论一致,我们也猜测蒜头果植株中可能存在植物-微生物共生的现象。

图 6 不同生境中蒜头果土壤理化性质(A)和土壤微生物生物量(B)的主成分分析(PCA)。
Figure 6. Principal components analysis (PCA) on physical and chemical properties (A) and soil microbial biomass (B) of Malania oleifera in idifferent habitats.
-
蒜头果的生长发育受到多种因素的影响,从土壤理化性质和土壤微生物来看,石山的蒜头果油含量和神经酸含量显著高于土山,可能是因为土山土壤pH、K、Ca、Mg、Fe、Zn、Si和真菌相对紧缺,导致石山与土山的头果油含量和神经酸含量有差异。因此,在保护野生蒜头果现有资源以及提高神经酸含量品质中,对土山的酸性土壤进行改善,并施加K、Ca、Mg、Fe、Zn肥和真菌微生物肥有利于提高蒜头果油含量和神经酸含量及品质,这些研究结果可为后续的保护和管理栽培高产量高品质的蒜头果植物提供科学参考。
致谢 感谢中国科学院西双版纳热带植物园公共技术中心对指标测定提供的支持。
Characterization of soil elements and microorganisms in Malania oleifera under different site conditions
More Information-
摘要: 不同居群蒜头果种子油及神经酸含量存在显著差异,为了探究土壤营养元素和微生物类型与蒜头果种子油含量及神经酸含量的关系,对蒜头果主要分布区不同居群的土壤理化性质和微生物组成进行了检测分析。结果表明,居群间土壤pH值、大量元素、中量元素、微量元素均存在显著差异,且与生境中岩石裸露率有一定的相关性,岩石裸露率高的石山的pH值显著高于岩石裸露率低的土山;除大量元素的P、S、Cu元素和NH4+-N、NO3−-N、AP的含量外,其他元素含量均为石山生境显著高于土山生境;石山生境的AMF、SF、Fungi生物量显著高于土山生境,而土山生境的Si含量显著高于石山生境;蒜头果土壤理化性质与土壤微生物之间存在一定的相关性,土壤Si含量与GP: GN显著正相关,但与pH值及其他元素含量呈显著负相关(P < 0.05);石山土壤pH值、K、Ca、Mg、Fe、Zn元素以及AMF、SF、Fungi含量高可能与石山生境中蒜头果种子油含量和神经酸含量高于土山生境有关,导致石山与土山的头果油含量和神经酸含量有差异。Abstract: There were significant differences in the seed oil and neuronic acid (NA) contents of Malania oleifera in different habitats. In order to investigate the relationship between soil nutrient elements and microbial types and the seed oil content and NA content of M. oleifera, soil physicochemical properties and microbial compositions of different habitats of M. oleifera in the main distribution areas were tested and analyzed. The results showed that there were significant differences in soil pH, massive elements, intermediate elements and trace elements among the habitats, and there was a certain correlation with the rate of rock exposure in the habitats, and the pH of the rocky mountains with a high rate of rock exposure was significantly higher than that of the dirt mountains with a low rate of rock exposure; except for the contents of P, S, Cu elements of massive elements, and the contents of NH4+-N, NO3−-N and AP, the contents of all other elements of the rocky mountains habitats were were significantly higher than those of the dirt mountains habitat; the biomass of AMF, SF, and Fungi was significantly higher in the rocky mountains habitat than in the dirt mountains habitat, whereas the content of Si was significantly higher in the dirt mountains habitat than in the rocky mountains habitat; there was a correlation between the physical and chemical properties of the soil and soil microorganisms, and the content of Si in the soil was significantly positively correlated with GP: GN but significantly negatively correlated with pH and the content of the other elements (P < 0.05); the rocky mountains High soil pH, K, Ca, Mg, Fe, Zn elements, and AMF, SF, Fungi contents may be related to the fact that the oil content of M. Oleifera and NA content were higher in the rocky mountains habitat than in the dirt mountains habitat, which led to the difference in the head fruit oil content and NA content between rocky mountains and dirt mountains.
-
Key words:
- habitats;
- elements;
- soil microorganism;
- PLFA;
- Malania oleifera
-
图 1 不同居群的蒜头果土壤理化性质(A)以及土壤微生物生物量(B)的差异(均值 ± SE)。不同小写字母表示蒜头果居群间的差异(P < 0.05)。
Fig. 1 Physical and chemical properties of the soil (A) and soil microbial biomass (B) of Malania oleifera in different populations (Mean ± SE). Different lowercase letters indicate significant Malania oleifera difference populations (P < 0.05).
图 2 不同居群间蒜头果土壤理化性质和土壤微生物生物量的 Spearman 相关性分析。注:*P < 0.05;**P < 0.01;***P < 0.001。
Fig. 2 Results on bivariate relationships on physical and chemical properties and soil microbial biomass of Malania oleifera in idifferent populations using Spearman correlation. Note: *P < 0.05; **P < 0.01; ***P < 0.001.
图 3 不同居群蒜头果土壤理化性质(A)和土壤微生物生物量(B)的聚类分析。
Fig. 3 Cluster analysis on physical and chemical properties (A) and soil microbial biomass (B) of Malania oleifera in idifferent populations.
图 4 不同生境的蒜头果土壤理化性质(A)以及土壤微生物生物量(B)的差异(均值 ± SE)。不同小写字母表示蒜头果生境间的差异(P < 0.05)。
Fig. 4 Physical and chemical properties of the soil (A) and soil microbial biomass (B) of Malania oleifera in different habitats (Mean ± SE). Different lowercase letters indicate significant Malania oleifera difference habitats (P < 0.05).
图 5 不同生境间蒜头果土壤理化性质和土壤微生物生物量的Spearman相关性分析。注:*P < 0.05;**P < 0.01;***P < 0.001。
Fig. 5 Results on bivariate relationships on physical and chemical properties and soil microbial biomass of Malania oleifera in idifferent habitats using Spearman correlation. Note: *P < 0.05; **P < 0.01; ***P < 0.001.
图 6 不同生境中蒜头果土壤理化性质(A)和土壤微生物生物量(B)的主成分分析(PCA)。
Fig. 6 Principal components analysis (PCA) on physical and chemical properties (A) and soil microbial biomass (B) of Malania oleifera in idifferent habitats.
表 1 样地概况
Tab. 1 Basic status of plots
地点 经度 纬度 海拔/m 生境 (岩石裸露率) 样本数 平均胸径(cm) 平均树高(m) 旧莫乡(JM) 104°90'E 23°92'N 1278.25 石山 (90%) 3 30.6 7.5 八宝镇(BB) 105°43'E 23°86'N 1027.49 石山 (70%) 3 21.8 8.5 曙光乡(SG) 105°16'E 23°72'N 1366.18 石山 (80%) 3 27.8 8.7 者桑乡(ZS) 106°08'E 23°74'N 444.04 土山 (5%) 3 20.7 12.8 板仑乡(BL) 105°73'E 23°65'N 843.00 土山 (5%) 3 24.9 9.5  下载: 导出CSV
下载: 导出CSV
-
[1] 李洪潮,常征,高明菊,等. 蒜头果研究进展[J]. 安徽农业科学,2020,48(17):8−10+14. [2] 李江莹. 几种含神经酸植物的种子脂肪酸组成与基因组特征研究[D]. 中国科学院大学,2020. [3] 蒲定福,冯自伟,郑仁健,等. 神经酸来源新方向的探讨[J]. 中国油脂,2021,46(8):76−80. [4] 赖家业. 珍稀植物蒜头果保护生物学研究[D]. 四川大学,2006. [5] 谢伟东,陈建华,赖家业,等. 蒜头果地理分布与水热关系分析[J]. 热带亚热带植物学报,2009,17(4):388−394. [6] 李小方,曹建华,徐祥明. GC-MS技术对比研究两种土壤上蒜头果果油品质[J]. 河南师范大学学报(自然科学版),2006(3):128−131. [7] HE X , LU T , LI J Y , et al. Germplasm resources of three wood plant species enriched with nervonic acid[J]. Plant diversity, 2022, 44(3): 308−315. [8] BOUAIN N; KORTE A; SATBHAI S B, et al. Systems genomics approaches provide new insights into Arabidopsis thaliana root growth regulation under combinatorial mineral nutrient limitation[J]. Plos genetics, 2019, 16(9): e1009107. [9] JACOBY R; PEUKEERT M; SUCCURRO A, et al. The Role of Soil Microorganisms in Plant Mineral Nutrition-Current Knowledge and Future Directions[J]. Frontiers in plant science, 2017(3): 1617. [10] 王茜,赵筱青,普军伟,等. 滇东南喀斯特区域石漠化时空格局演变研究-以广南县为例[J]. 中国岩溶,2021,40(4):707−717. [11] 苗培培,赵筱青,普军伟,等. 喀斯特山区生态系统服务权衡/协同时空分异研究-以云南广南县为例[J]. 山地学报,2023,41(1):103−114. [12] 袁鸿文. 富宁县油茶产业现状及发展对策[J]. 林业调查规划,2014,34(3):76−79. [13] DEVIN L W, TERI C B. Toward conceptual clarity: PLFA in warmed soils[J]. Soil biology and biochemistry, 2013, 57: 769−774. doi: 10.1016/j.soilbio.2012.08.016 [14] RAINER G J. Phospholipid fatty acids in soil-drawbacks and future prospects[J]. Biology and fertility of soils, 2022, 58: 1−6. doi: 10.1007/s00374-021-01613-w [15] 赖家业,杨振德,文祥凤. 两种立地条件下蒜头果叶绿素含量比较研究[J]. 广西植物,1999(3):272−276. [16] 石松利,王迎春,周健华. 不同生境条件四合木(Tetraena mongolica)内源激素季节变化及种群差异[J]. 生态学报,2009,29(5):2252−2262. [17] 朱庆征,冯志培,冯二朋,等. 祁连山区不同海拔植被带土壤微生物磷脂脂肪酸分析[J/OL]. 中国水土保持科学(中英文),2023:1−13. [18] 何越,李春涛,俞元春,等. 亚热带森林土壤微生物生物量及群落功能特征的城乡梯度变化[J]. 应用生态学报,2021,32(1):93−102. [19] 郭川,周冀琼,张英俊. 宿主和养分介导了AMF和根瘤菌对植物生长的协同效应[J]. 草地学报,2023,31(7):1931−1938. [20] DAVIDE F , SOPHIE Q V R, JASPER V R , et al. Plant functional group drives the community structure of saprophytic fungi in a grassland biodiversity experiment [J]. Plant soil, 2021, 461: 91-105. [21] KANG P, PAN Y Q , RAN Y C, et al. Soil saprophytic fungi could be used as an important ecological indicator for land management in desert steppe [J]. Ecological indicators, 2023, 150 : 110224. [22] WANG Z, DENG X Z, SONG W, et al. What is the main cause of grassland degradation? A case study of grassland ecosystem service in the middlesouth Inner Mongolia. Catena, 2017, 150: 100–107. [23] 沈清清,刘芳,胡彦. 药用植物根腐病病原菌研究进展[J]. 北方园艺,2014(11):187−190. [24] 席守鸿,明安刚,谭玲,等. 南亚热带乡土树种与桉树人工林土壤真菌群落多样性和功能类群的比较[J/OL]. 广西植物,2023:1-15. [25] 刘炜璇,李依蒙,江红星,等. 吉林莫莫格国家级自然保护区四种典型植物群落下土壤微生物组成的对比分析[J/OL]. 生态学杂志,2023:1−12. [26] 李佳奇,郭屹立,李冬兴,等. 桂西南北热带喀斯特季节性雨林土壤钾、钙、镁空间分布特征及其影响因素[J]. 生物多样性,2023,31(2):178−189. [27] TUBANA B S; BABU T and DATNOFF L E. A review of silicon in soils and plants and its role in US agriculture: history and future perspectives[J]. Soil Science, 2016, 181(9/10): 393−411. doi: 10.1097/SS.0000000000000179 [28] GRAŠIČ M, SAKOVIČ T, ABRAM D, et al. Do soil and leaf silicon content affect leaf functional traits in Deschampsia caespitosa from different habitats?[J]. Biologia plantarum, 2020, 64: 234−243. doi: 10.32615/bp.2019.155 [29] 敬洪霞,孙宁骁,UMAIR M,等. 滇南喀斯特地区不同季节土壤和灌木叶片化学计量特征及对水分添加的响应[J]. 植物生态学报,2020,44(1):56−69. [30] 吴求生,龙健,李娟,等. 茂兰喀斯特森林小生境类型对土壤微生物群落组成的影响[J]. 生态学报,2019,39(3):1009−1018. [31] 邓杰文,石杨,李斌,等. 微生物在沙化土壤修复中的应用研究进展[J]. 应用与环境生物学报,2022,28(5):1367−1374. [32] 张仲富,王四海,杨卫,等. 蒜头果根际细菌群落结构与功能特征对其健康状态的响应[J]. 植物生态学报,2023,47(7):1020−1031. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 12
- HTML全文浏览量: 5
- PDF下载量: 0
- 被引次数: 0









