






 下载:
下载:


-
苔藓植物作为自然演替过程中常见的先锋物种,其独特的生理结构和脱水复苏能力使其能在裸岩、沙漠、极地等诸多极端恶劣环境下仍具有广泛的适应性和强大的繁殖力[1],在水源涵养、土壤保持、生物多样性维持等生态系统服务效益等方面起着重要的调节作用。生物土壤结皮(biological soil crusts, BSCs)又称生物结皮,是由不同种类的藻类、地衣、苔藓以及微生物等类群与土壤表层颗粒有机结合形成的复合体[2]。苔藓作为地表覆被物之一,往往通过菌丝体、假根和分泌物与表层土壤颗粒“胶结”参与形成生物结皮,在高寒、干旱半干旱地区等大多数生态系统中发挥着重要的生态功能[3,4]。有研究表明[4-6],苔藓可通过分泌有机酸、释放CO2和机械作用溶解岩面,积累丰富的有机质,具有成土功能,进而促进其它植物类群的萌发、定居和存活,并为许多土壤微生物和微小动物提供适宜的生境和食物来源。此外,苔藓还有很强的吸水能力,能减缓直立峭壁、高陡边坡径流量和径流速度,发挥着水源涵养、水土保持功能等重要的生态功能[7,8]。
九寨沟国家自然保护区成立于1978年,是我国第一个以保护自然风景为主要目标的国家级自然保护区,先后入选《世界自然遗产名录》、国际“人与生物圈”保护网络。然而在2017年九寨沟保护区内发生的7.0级地震致使当地山体裸露,引发了大量崩塌、滑坡、泥石流等次生灾害,地表植被覆盖率及生物多样性减少,动物栖息地受到不同程度影响,生态环境质量受到极大挑战[9-11],亟待开展九寨沟震灾后生态恢复与重建工作。前人研究结果表明,在九寨沟裸岩边坡植被修复的大量实例中,原生草种、商业草种、乔木、灌木是最常见的修复方法[12]。然而考虑到九寨沟震后少基质,甚至无基质的裸岩环境,传统的植树种草等恢复措施可能并不能被视为最理想的修复方法[12,13]。现有大量研究发现苔藓植物的大面积生长对生态脆弱区生态恢复效益的影响显著,受到国内外相关学者的广泛关注[14-17]。因此,研究基于当地不同生境下东亚砂藓(Racomitrium japonicum)、大灰藓(Hypnum plumaeforme)、与阔边匐灯藓(Plagiomnium ellipticum)3种典型苔藓植物,比较它们溶蚀作用、固土以及保水能力的生态恢复效应,筛选出抗逆性好、水土涵养功能强的特定藓种,以加快推进震灾造成的九寨沟大面积裸露、水土流失等问题的生态修复工作,对保障九寨沟景观美学价值、生态安全,构筑生态屏障具有重要现实意义。
-
九寨沟国家自然保护区地处青藏高原东缘,位于四川省阿坝藏族羌族自治州九寨沟县漳扎镇境内(东经103°46′14″~104°5′3″、北纬32°54′13″~33°19′57″)。九寨沟海拔1000—4500 m,年均温为6-7℃,月平均最低气温−3.7℃,月平均最高气温16.8℃,分别出现在1月和7月,年降雨量为696.6-957.5 mm[18,19],气候寒冷干燥,降水丰富,相对湿度较大。保护区内地势复杂,主要为喀斯特地形,存在大量岩石裸露,水分涵养能力较弱,属于我国西部典型的山地生态脆弱区[19]。由于区域地理位置的特殊性、气候条件与地形的复杂性,蕴藏着丰富的动植物资源,植被覆盖率超过80%,其中苔藓植物物种丰富,代表种有大羽藓(Thuidium cymbifolium)、大灰藓(Hypnum plumaeforme)、银藓(Anomobryum filiforme)等[19]。
-
基于文献查阅与前期对九寨沟内苔藓种类的实地调查[19-21],选择东亚砂藓、大灰藓与阔边匍灯藓三个广布种作为研究对象(见图1)。2019年9月在四川省九寨沟国家自然保护区选取苔藓结皮4种典型的生境类型(草地、灌木、乔木林以及空地),每个生境类型下布设3个1 m×1 m 的样方,在每个样方内按5点取样法将采集的苔藓结皮样品混合均匀,重复采集3次。保证采集时将结皮轻轻揭起并保留结皮下方土壤(结皮表下5 mm左右),每块结皮的面积不小于10×10cm2。

图 1 东亚砂藓(a)、大灰藓(b)与阔边匍灯藓(c)野外生境图
Figure 1. Different habitat conditions of Racomitrium japonicum (a), Hypnum plumaeforme (b), and Plagiomnium ellipticum (c)
-
参照前人[22,23]的研究方法略作改动以测定植物碳酸酐酶(Carbonic Anhydrase, CA)活性。称取0.3-0.4 g植物样品在研钵中加入液氮充分研磨,再加入5 mL巴比妥缓冲液(20 mmol·L−1, pH=8.3)混合,于16000 r·min−1下离心15 min,取上清液0.5 mL,加入到含10 mL巴比妥-KOH缓冲液(pH=8.3)的反应室中并搅拌均匀。再将pH计置于反应液中,记下起始的pH值后迅速加入10 mL冰冻CO2饱和水溶液,用秒表计时,待pH值下降一个单位时,停止计时,记下反应时间为Te。并用同一样品的煮沸液做对照,记下反应时间为To。CA活性以每克叶片含有的酶活单位数(U·g−1 DW)表示,其中U=10(To/Te-1)。此外,根据曾成[24]等的溶蚀试片法估算苔藓溶蚀岩面的溶蚀速率与碳汇量。
-
将苔藓结皮层与土壤层彻底分离,将洗净风干后的植物样品置于烘箱中,在60℃的条件下烘48 h后测量干重。土壤样品在室温下风干24 h后过60目分样筛,称量其土壤总重,即为苔藓固土量。固土率是苔藓固土能力的直观表现。计算公式如下:
$$ SP=SW/PDW \times 100 \text% $$ (1) 式中:SP为固土率(%),SW为固土量(kg·hm−2),PDW为植物样品干重(kg·hm−2)。
-
参考前人的方法测定苔藓植物的保水量和保水率[25,26],研究采用室内浸泡法进行测定。将苔藓植物在水中浸泡24 h后,将苔藓植物样品置于纱网上,待滴干重力水后称其湿重,在60℃恒温箱内干燥48 h至恒重,称量其干重。计算公式如下:
$$ W=(PEW-PDW)/PDW\times 100 \text% $$ (2) 式中:W为保水率(%),PFW为植物样品湿重(kg·hm−2),PDW为植物样品干重(kg·hm−2)。
-
所测样品均重复3次试验并取平均值,利用Excel 2016和SPSS 22.0进行统计分析与数据处理,结合单因素方差分析(One-way ANOVA) 与最小差异法(LSD)检验数据间的显著性差异(P<0.05),采用SigmaPlot 10.0制图。文中数据用平均值(Mean)±标准差(SD)表示。
-
不同苔藓物种的CA活性、溶蚀速率、碳汇量均存在差异(表1),其中东亚砂藓具有最高的CA活性,进而具备最大的溶蚀速率(4.8×10−4 mg·m−2·a−1)与碳汇量(2.12 t·km−2·a−1 CO2),远高于大灰藓和阔边匍灯藓。结果表明,东亚砂藓保持较强的酶促溶蚀驱动作用和溶岩能力,适宜在石灰岩生长。
表 1 三种苔藓CA活性、溶蚀速率与碳汇量估算
Table 1. CA activity, estimation of dissolution rate and carbon sink of three bryophytes
苔藓种类 CA活性
(U·g−1 DW)溶蚀速率
(t·km−2·a−1CO2)碳汇量估算
(mg·m−2·a−1)东亚砂藓 50.5±2.5 4.8×10−4 2.12 大灰藓 30.7±5.4 2.6×10−4 1.15 阔边匍灯藓 12.5±1.8 1.5×10−4 0.59 -
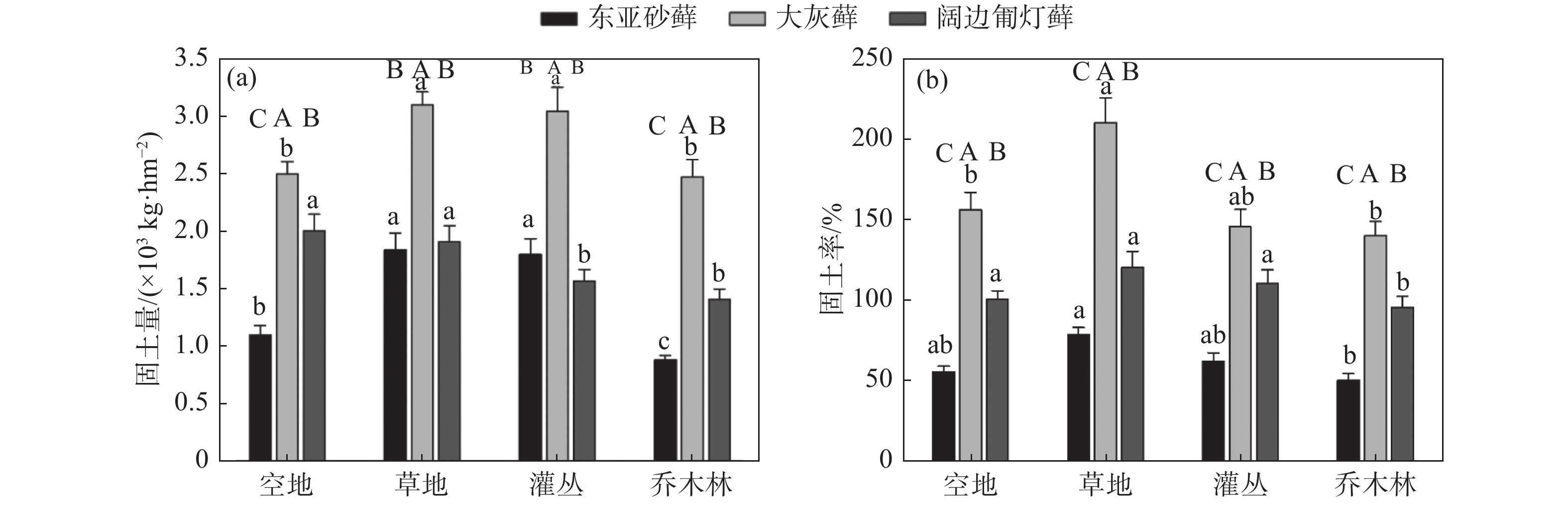
通过对样品固土量对比发现,不同生境类型下苔藓结皮的固土量不同,且不同苔藓植物的固土量具有差异。研究区域内生于草地的苔藓植物固土量相对高于其它生境,此外,大灰藓在4种生境类型下的固土量较高,尤其是草地中大灰藓的固土量最高,达到了3.1×103 kg·hm−2(见图2a),最高固土率也为自身干重的2倍多(见图2b)。对东亚砂藓而言,草地和灌丛生境下的固土量显著高于空地与乔木林两种生境(P<0.05),而阔边匍灯藓在空地与草地的生境下的固土量的差异不显著(P>0.05),但显著高于另外两种生境(P<0.05)。
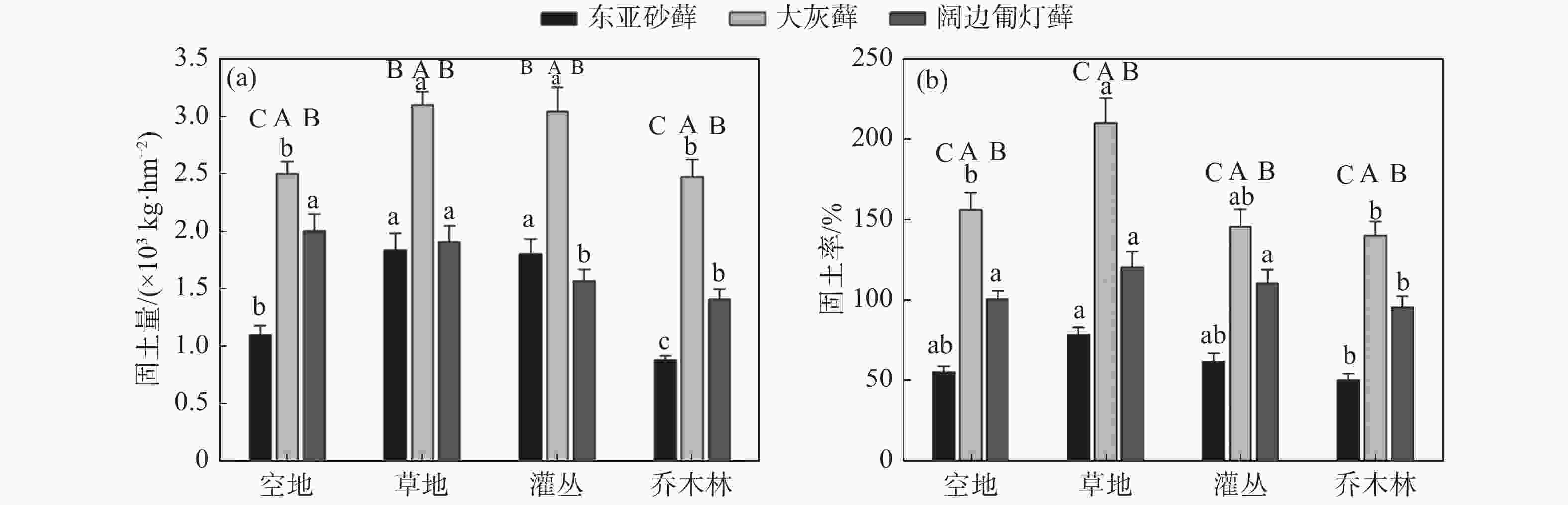
图 2 不同生境条件下三种苔藓的固土量(a)与固土率(b)比较
Figure 2. Comparison of soil consolidation capacity (a) and rate (b) of three bryophytes under different habitat types
3种苔藓植物的固土率变化趋势与固土量趋势大致相同,表现为草地生境下3种苔藓植物的平均固土率较其它生境仍然较高,总体表现为草地>灌丛>空地>乔木林;而对于同一生境下的苔藓植物而言,大灰藓的固土率仍然最高,阔边匍灯藓次之,东亚砂藓中最低(见图2b)。
-
比较相同苔藓物种在不同生境下的保水量差异并不显著(P>0.05),然而空地上的平均持水量表现略高,具体呈现出空地>草地>灌丛>乔木林的趋势(见图3a)。不同生境条件下3种苔藓的保水量显著不同(P<0.05),表现为各生境中阔边匍灯藓的保水量最高(>1100 kg·hm−2),其次为大灰藓(1000-1200kg·hm−2),东亚砂藓的保水量最低(<1000 kg·hm−2)(见图3a)。
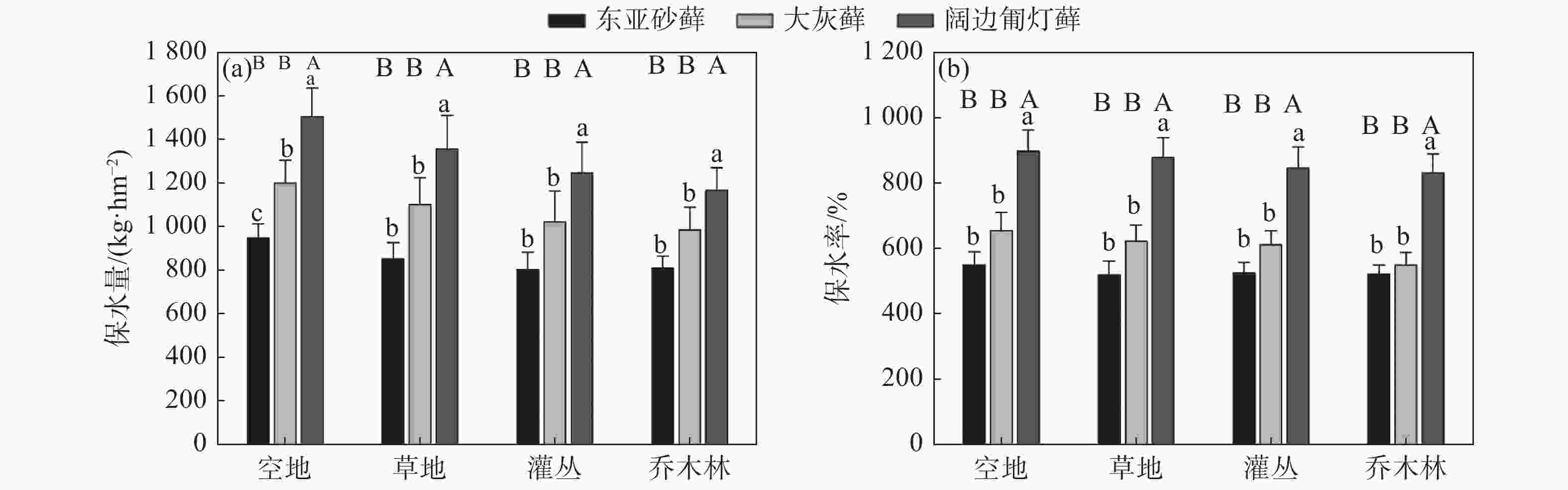
图 3 不同生境条件下三种苔藓的保水量(a)与保水率(b)比较
Figure 3. Comparison of water holding capacity (a) and rate (b) of three bryophytes under different habitat types
不同苔藓植物的保水率变化趋势与保水量一致(见图3b),相同苔藓物种的保水率在不同生境下的差异不大(P>0.05),而对于不同藓种而言,阔边匍灯藓的保水率仍然显著高于灰藓与砂藓(P<0.05),其在各生境下的保水率均可达到自身干重的8倍以上,大灰藓与东砂藓则能达到自身干重的5倍以上(见图3b)。
-
碳酸酐酶是植物中普遍存在的一种含锌金属酶,不仅在植物的光合作用起着重要作用[22,27],而且能作为生物催化剂决定碳酸盐岩的溶解速率,促进岩石的风化、土壤的形成和发育[28]。一般而言,生长在岩面、岩壁和岩面薄土等贫瘠地区上的苔藓植物CA活性较高[29]。研究通过测定不同生境下东亚砂藓、大灰藓与阔边匍灯藓3种藓种的碳酸酐酶活性、溶蚀速率和碳汇量,发现3种苔藓植物均能检测到不同程度的CA活性,这说明CA在不同生境的苔藓植物体中广泛存在,这与已有的研究结果相同[28,29]。然而不同的藓种之间具有明显差异,表现为东亚砂藓(50.5 U·g−1 DW)>大灰藓(30.7 U·g−1 DW)>阔边匍灯藓(12.5 U·g−1 DW),东亚砂藓同时具有最大的酶促溶蚀速率(4.8×10−4 mg·m−2·a−1) (见表1),远高于大灰藓和阔边匍灯藓,推测这可能与植物本身的形态结构、生理生态以及土壤环境有关,其内在的相关机制有待进一步深入研究。此外,有研究发现苔藓植物的岩溶作用对大气中CO2也具有显著的捕获回收效应[30,31]。研究中发现3种苔藓岩溶过程中产生的碳汇量有所不同,这在一定程度上为植物的固碳增汇提供基础的参考数据。
-
九寨沟地震导致山体大量的岩石崩塌、滑坡,形成了大量巨大裸露石质岩面,土层浅薄,水土资源匮乏,大部分维管束植物在此类生境中生存极为困难。而苔藓作为最原始的高等植物,能较好适应裸岩、干燥的极端环境,这往往离不开苔藓植物的成土特性与较强的水土保持能力[4-6]。研究结果表明,3种苔藓植物均具有较强的固土能力,最高固土量为3.1×103 kg·hm−2(见图2a),最高固土率可达到自身干重的2倍以上(见图2b)。3种苔藓植物之间的固土量、固土率差异显著。其中,大灰藓在各生境类型下均表现出最高的固土量、固土率,阔边匍灯藓次之,东亚砂藓中最低。可见,苔藓植物类型差异会较大影响自身的固土能力。究其原因在于大灰藓植株矮小,属于密集交织型,有利于对灰尘和土壤小颗粒的沉积与固定,而阔边匍灯藓植物叶边缘平滑,且植株蓬松,而东亚砂藓为丛生型,单株可独立生长,因而固土效率较低。相关报道指出,在没有外界的干扰下,苔藓植物茎叶与假根的发育对其固土能力起着关键性作用[32],提高假根与根长密度的比重,有利于加大苔藓植物的生物量,加速土壤的形成速率。因此不同苔藓植物的体现不同的固土能力依赖其独特的形态结构与生理特性。
研究区域内草地中苔藓植物的固土量相对高于其它生境下苔藓植物固土量,表明苔藓植物固土能力还受空间异质性和生境类型的影响。草地大多以低矮草本层为主,植被覆盖率较低,加之缺乏林灌层的保护,易遭受强风、降雨的对土壤的直接冲击,侵蚀过程中灰尘和土壤细小颗粒不断被带到岩石表面,最终由苔藓植物沉积固定[1,33]。应特别指出的是,后期当苔藓与表层土壤复合后形成的苔藓结皮,有利于增强表土的稳定性,延长沉积物在结皮层中的滞留时间,在一定程度上减缓了土壤免受风蚀和水蚀的能力[34]。
-
由于苔藓植物疏松多孔、表面强大、具有海绵性状的弹性力学特征,其假根和植物茎叶快速吸收水分,往往能够蓄存大量水分,在水分平衡中起到了涵养和调节的重要作用[7,8]。研究结果显示,各生境下不同苔藓植物的保水能力均高于植物体自身干重,且种间差异显著,尤其以阔边匍灯藓的持水能力最强,持水量维持在1100-1400 kg·hm−2之间(见图3a),持水率可达到自身干重的8倍以上(见图3b)。考虑到苔藓植物的生活型与生物量特征也许会影响植物体自身的吸水保水能力[35],推测阔边匍灯藓的持水量较高的原因可能是其匍匐茎纵横交错成片生长,有利于提高与水分的接触面积。
此外,在研究区内,相同苔藓物种在不同生境下的持水能力无显著差异,空地上的平均持水量表现略高,具体呈现出空地、草地、灌丛、乔木林逐渐降低的趋势,说明苔藓层在裸露的基岩表面生长具有一定的优势,这可能与其较低的初始含水量有关。这与李军峰等[26]人的研究结果相同。
综上,建议从成土、固土、保水和生态修复的角度出发,充分考虑东亚砂藓碳酸酐酶高,溶岩成土能力较强的特点,作为九寨沟喀斯特岩面生态恢复的先锋苔藓;中期利用大灰藓交织结皮固土能力强的优势,提高固土量,加快土壤的形成与累积;后期借助阔边匍灯藓吸水、保水能力,发挥海绵体的吸水储水能力作用,来主动调节水文生态过程。总之,在不同阶段、不同生境下选择特定藓种,充分发挥苔藓植物生态系统工程师的先锋作用,以推动九寨沟地区震后的生态恢复与重建。
Research on ecological effects of typical moss crusts in Jiuzhaigou National Nature Reserve
More Information-
摘要: 苔藓植物作为初生演替阶段的先锋种,其独特的生理结构和脱水复苏能力在水源涵养、土壤保持、生物多样性维持等方面发挥着重要作用。以不同生境下(草地、灌木、乔木林以及空地)东亚砂藓(Racomitrium japonicum)、大灰藓(Hypnum plumaeforme)及阔边匐灯藓(Plagiomnium ellipticum)3种苔藓植物为研究对象,通过野外实地考察与室内分析,评估3种典型苔藓结皮的溶蚀能力、固土以及保水特性对九寨沟国家自然保护区震后恢复的生态效应。结果表明:东亚砂藓具有最高的碳酸酐酶活性(50.5 U·g−1 DW),进而具备最大的溶蚀速率(4.8×10−4 t·km−2· a−1 CO2)与碳汇量(2.12 mg·m−2·a−1)。不同生境条件下的不同苔藓固土能力不同,其中大灰藓结皮层的固土量最高(3.1×103 kg·hm−2),最高固土率可达到自身干重的2倍以上。此外,阔边匍灯藓的保水能力均高于大灰藓与东亚砂藓,其保水量维持在1100-1400 kg·hm−2之间,保水率可达自身干重的8倍以上。综上,在利用苔藓结皮进行生态恢复过程中,建议从成土、固土、保水等生态修复的角度出发,针对不同阶段、不同生境条件选择特定藓种,以加快九寨沟地区震后的生态恢复与重建。
-
关键词:
- 苔藓植物;
- 苔藓结皮;
- 九寨沟国家自然保护区;
- 生态效应;
- 生态恢复
Abstract: As a pioneer species in the primary succession stage, bryophytes play an important role in water conservation, soil conservation and biodiversity maintenance due to their unique physiological structure and dehydration recovery ability. In this study, three bryophytes including Racomitrium japonicum, Hypnum plumaeforme and Plagiomnium ellipticum in different habitats (grassland, shrub, forest and bare rock) were selected as the research objects. Based on field investigation and laboratory analysis, the ecological effects of dissolution activity, soil and water holding capacities of three typical bryophytes crusts on the post-earthquake restoration in the Jiuzhaigou National Nature Reserve were evaluated. The results showed that R. japonicum had the highest the carbonic anhydrase (CA) activity (50.5 U·g−1 DW), the maximum dissolution rate (4.8×10−4 t·km−2·a−1 CO2) and carbon sink content (2.12 mg ·m−2·a−1). Different mosses in different habitats had different soil consolidation capacity, among which the soil consolidation capacity of H. plumaeforme was the highest (3.1×103 kg·hm−2), and its maximum soil consolidation rate was more than 2 times of its own dry weight. In addition, the water retention capacity of P. ellipticum was higher than that of R. japonicum and H. plumaeforme. The water holding capacity of P. ellipticum was between 1100-1400 kg·hm−2, and the water retention rate was more than 8 times of its own dry weight. In conclusion, in the process of ecological restoration by using moss crusts, it is suggested to select specific bryophytes according to different stages and different habitat conditions from the perspective of ecological restoration such as soil formation, soil consolidation and water conservation, so as to accelerate the ecological restoration and reconstruction after the earthquake in Jiuzhaigou National Nature Reserve. -
图 1 东亚砂藓(a)、大灰藓(b)与阔边匍灯藓(c)野外生境图
Fig. 1 Different habitat conditions of Racomitrium japonicum (a), Hypnum plumaeforme (b), and Plagiomnium ellipticum (c)
图 2 不同生境条件下三种苔藓的固土量(a)与固土率(b)比较
注:不同大写字母表示同种生境类型下不同苔藓植物之间的差异显著(P<0.05);不同小写字母表示同种苔藓植物不同生境类型下之间的差异显著(P<0.05),下同。
Fig. 2 Comparison of soil consolidation capacity (a) and rate (b) of three bryophytes under different habitat types
图 3 不同生境条件下三种苔藓的保水量(a)与保水率(b)比较
Fig. 3 Comparison of water holding capacity (a) and rate (b) of three bryophytes under different habitat types
表 1 三种苔藓CA活性、溶蚀速率与碳汇量估算
Tab. 1 CA activity, estimation of dissolution rate and carbon sink of three bryophytes
苔藓种类 CA活性
(U·g−1 DW)溶蚀速率
(t·km−2·a−1CO2)碳汇量估算
(mg·m−2·a−1)东亚砂藓 50.5±2.5 4.8×10−4 2.12 大灰藓 30.7±5.4 2.6×10−4 1.15 阔边匍灯藓 12.5±1.8 1.5×10−4 0.59  下载: 导出CSV
下载: 导出CSV
-
[1] 吴玉环,程国栋,高谦. 苔藓植物的生态功能及在植被恢复与重建中的作用[J]. 中国沙漠,2003,23(3):9−14. [2] Nevins C J, Inglett P W, Strauss S L. Biological soil crusts structure the subsurface microbiome in a sandy agroecosystem[J]. Plant and Soil, 2021, 462(1): 311−329. [3] Oguri E, Deguchi H. Radiocesium contamination of the moss <italic>Hypnum plumaeforme</italic> caused by the Fukushima Dai-ichi Nuclear Power Plant accident[J]. Journal of environmental radioactivity, 2018, 192: 648−653. doi: 10.1016/j.jenvrad.2018.02.013 [4] 许欢欢,张宝琦,汪建芳,等. 黄土高原典型生物结皮对坡面产流产沙过程的影响[J]. 水土保持通报,2020,40(6):8−13. [5] Kinnell P I A, Chartres C J, Watson C L. The effects of fire on the soil in a degraded semiarid woodland. II. Susceptibility of the soil to erosion by shallow rain impacted flow[J]. Soil Research, 1990, 28(5): 779−794. doi: 10.1071/SR9900779 [6] Belnap J, Welter J R, Grimm N B, et al. Linkages between microbial and hydrologic processes in arid and semiarid watersheds[J]. Ecology, 2005, 86(2): 298−307. doi: 10.1890/03-0567 [7] Wang B, Wu F Z, Xiao S, et al. Effect of succession gaps on the understory water-holding capacity in an over-mature alpine forest at the upper reaches of the Yangtze River[J]. Hydrological Processes, 2016, 30(5): 692−703. doi: 10.1002/hyp.10613 [8] 从春蕾,刘天雷,孔祥远,等. 贵州普定喀斯特受损生态系统石生藓类植物区系及物种多样性研究[J]. 中国岩溶,2017,36(2):179−186. [9] Wang J ,Jin W ,Cui Y , et al.Earthquake-triggered landslides affecting a UNESCO Natural Site: the 2017 Jiuzhaigou Earthquake in the World National Park, China[J].Journal of Mountain Science,2018,15(7):1412−1428. [10] 李孝永,杜国明,匡文慧. 九寨沟地震次生灾害风险及对自然保护区和自然遗产地的影响[J]. 水土保持通报,2019,39(2):301−308. [11] Ling S ,Sun C ,Li X , et al.Characterizing the distribution pattern and geologic and geomorphic controls on earthquake-triggered landslide occurrence during the 2017 Ms 7.0 Jiuzhaigou earthquake, Sichuan, China[J].Landslides,2021,18:1275−1291. [12] 陈云,董发勤,韩颖,等.九寨沟景区“8·8”地震前后及钙华生境保育修复后植被和水系的动态变化[J].中国岩溶,2021,40(01):68−76. [13] 孙俊峰,陈其兵,王怡,等. 苔藓植物联合乡土草种应用于植被恢复工程的初步研究[J]. 四川草原,2005(3):16−18. [14] 左元彬,辜彬,艾应伟. 苔藓植物在道路边坡植被生态恢复中的应用与分析[J]. 中国水土保持科学,2006,4(s1):122−125. [15] Song L, Zhang Y J, Chen X, et al. Water relations and gas exchange of fan bryophytes and their adaptations to microhabitats in an Asian subtropical montane cloud forest[J]. Journal of Plant Research, 2015, 128(4): 573−584. doi: 10.1007/s10265-015-0721-z [16] 刘鑫,包维楷,胡斌,等.高寒山区道路边坡植被恢复物种选择及适宜性评估[J].应用与环境生物学报,2016,22(06):1015−1022. [17] Bu C, Li R, Wang C, et al. Successful field cultivation of moss biocrusts on disturbed soil surfaces in the short term[J].Plant and Soil, 2018, 429(1-2): 227−240. [18] 张炜,代金莉,贺维,等. 九寨沟地震灾后植被恢复过程中树种筛选[J]. 四川林业科技,2020,41(6):117−123. [19] 夏红霞,朱大林,张跃,等. 九寨沟国家级自然保护区藓类植物多样性及地理区系[J]. 应用与环境生物学报,2022,2 8(6):1615-1621. [20] Lei Y B, Xia H X, Chen K, et al. Photosynthetic regulation in response to fluctuating light conditions under temperature stress in three mosses with different light requirements[J]. Plant Science, 2021, 311: 111020. doi: 10.1016/j.plantsci.2021.111020 [21] 孙庚,类延宝,陈珂,等. 九寨沟国家级自然保护区苔藓植物图鉴[M],成都:四川科学技术出版社,2021. [22] Brownell P F, Bielig L M, Grof C P L. Increased carbonic anhydrase activity in leaves of sodium-deficient C<sub>4</sub> plants[J]. Functional Plant Biology, 1991, 18(6): 589−592. doi: 10.1071/PP9910589 [23] 郭敏亮,高煜珠,王忠. 用酸度计测定植物碳酸酐酶活性[J]. 植物生理学通讯,1988(6):59−61. [24] 曾成,赵敏,杨睿,等. 岩溶作用碳汇强度计算的溶蚀试片法和水化学径流法比较—以陈旗岩溶泉域为例[J]. 水文地质工程地质,2014,41(1):106−111. [25] 徐杰,白学良,杨持,等. 固定沙丘结皮层藓类植物多样性及固沙作用研究[J]. 植物生态学报,2003(4):545−551. [26] 李军峰,王智慧,张朝晖. 喀斯特石漠化山区苔藓多样性及水土保持研究[J]. 环境科学研究,2013,26(7):759−764. [27] 刘再华. 碳酸酐酶对碳酸盐岩溶解的催化作用及其在大气CO<sub>2</sub>沉降中的意义[J]. 地球学报,2001(5):477−480. [28] 张楷燕,李同建,张显强,等. 3种石生苔藓植物碳酸酐酶对石灰岩的溶蚀作用[J]. 中国岩溶,2017,36(4):441−446. doi: 10.11932/karst20170403 [29] 刘天雷,从春蕾,胡丹,等. 贵州普定6种喀斯特石生植物及其土壤的碳酸酐酶活性[J]. 中国岩溶,2017,36(2):187−192. doi: 10.11932/karst20170205 [30] Zhong C J, Dao X Y. CO<sub>2</sub> source-sink in karst processes in karst areas of China[J]. Episodes Journal of International Geoscience, 1999, 22(1): 33−35. [31] Sancho L G, Belnap J, Colesie C, et al. Carbon budgets of biological soil crusts at micro-, meso-, and global scales[M]//Biological soil crusts: an organizing principle in drylands. Berlin: Springer, 2016: 287−304. [32] 刘润,申家琛,张朝晖. 4种苔藓植物在喀斯特石漠化地区的生态修复意义[J]. 水土保持学报,2018,32(6):141−148. [33] 赵海燕,郭荣慧,段宇涛,等. 苔藓植物护坡及固土作用试验研究[J]. 兰州大学学报(自然科学版),2018,54(3):364−368. [34] 张冠华,易亮,丁文峰,等. 三峡库区苔藓生物结皮对土壤水分入渗的影响[J]. 应用生态学报,2022,33(7):1835−1842. doi: 10.13287/j.1001-9332.202207.001 [35] 涂娜,严友进,戴全厚,等. 喀斯特石漠化区典型生境下石生苔藓的固土持水作用[J]. 生态学报,2021,41(15):6203−6214. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 112
- HTML全文浏览量: 42
- PDF下载量: 12
- 被引次数: 0











