






 下载:
下载:

-
成都麻羊,又名四川铜羊,原产于成都平原及附近丘陵地区,是我国优良肉用地方山羊品种[1, 2]。成都麻羊具有生长发育快、体格较大、性早熟、繁殖能力强、屠宰率高、膻味轻、肉质嫩、板皮优良、肉皮兼用等显著特点,现有丘陵型和山地型两个生态类型[1-3]。在我国,早有引进成都麻羊改良当地山羊培育新品种的案例,如四川省有名的南江黄羊、金堂黑山羊品种[1-3]。但是,圈养规模较小、缺乏科学的管理、近亲繁殖现象等问题会导致成都麻羊的遗传变异丢失[4],引起圈养种群退化,长期会导致遗传多样性逐渐丧失,一些特有的重要基因资源会在培育中消失殆尽,同时会造成个体适应能力变弱,抗病力差,死亡率增加等[5],长期下来会影响圈养成都麻羊种群的健康发展。畜禽遗传资源本质上是基因资源,保护畜禽遗传资源就是保护基因的多样性[6]。因此,成都麻羊作为我国山羊品种的一个宝贵基因库,维持其种群遗传多样性,最大限度地降低遗传损失,保证其种群遗传健康,对其种群种质资源的良性发展和种群的遗传管理都具有重要意义。
微卫星DNA(microsatellite DNA),是以少数几个核苷酸为基本单位进行多次串联重复的DNA序列,因而也被称为简单序列重复(simple sequence repeat,SSR)或短串联重复序列(short tandem repeat,STR)[6-9]。相比其他的分子标记技术,微卫星DNA标记优点突出,如呈现共显性遗传、高度多态性等,是目前最为准确、先进的遗传标记系统之一,是种群遗传结构研究和多样性评估最常用的工具[5, 7, 10-14]。在山羊、绵羊中,利用微卫星DNA标记测定遗传多样性已有较多相关研究[9, 11, 15-21]。
鉴于此,为了加强成都麻羊资源的保护和利用,实现优质地方山羊品种种质资源的良性开发,满足市场对成都麻羊的发展需求,借助微卫星DNA作为遗传标记,对成都麻羊圈养种群进行遗传多样性现状调查,以期为该资源群体的遗传背景研究提供分子理论依据,同时对合理利用成都麻羊资源和可持续发展也具有重要意义。
-
本研究以成都西岭雪农业开发有限公司(四川省成都市大邑县)饲养的成都麻羊为研究对象,共随机挑选268只个体采集血液样品,其中雄性个体18只,雌性个体250只。对每只成都麻羊个体进行颈静脉采血5 mL,随后用EDTA抗凝混匀,置于冰盒中带回实验室,−20 ℃保存。
-
本研究采用Tianamp基因组DNA提取试剂盒提取DNA,提取步骤参照使用说明书执行。将提取得到的DNA用1%的琼脂糖凝胶进行电泳检测,并将其保存于−20 ℃冰箱。
参考已发表文献[7, 9, 10, 16, 17, 20, 21],共选46对多态性微卫星引物委托生工生物工程(上海)股份有限公司合成。通过预实验筛选,有9对引物(见表1)能较稳定地扩增得到多态性较高的微卫星位点。PCR扩增体系(20 uL):DNA模板(浓度25 ng/uL)2 uL,正向引物(浓度10 umol/ul)0.5 uL,反向引物(浓度10 umol/ul)0.5 uL,Mix溶液10 uL,灭菌双蒸水7 uL。PCR扩增条件:首先95 ℃预变性持续15 min;之后94 ℃变性持续30 s,相应退火温度退火持续30 s,72 ℃延伸持续30 s,并设置30个循环;最后72 ℃终止延伸持续10 min。每次扩增过程中均设定阴性对照,每个样品扩增3次。PCR扩增产物经1.5%的琼脂糖凝胶电泳检测后拍照。
表 1 本研究所用微卫星DNA标记
Table 1. Microsatellite DNA markers were used in this study
微卫星位点 引物序列(5′-3′) 5′端修饰 退火温度/℃ ILSTS004 F: CTTAAAATCTGTCTTTCTTCC 5`6-FAM 48 R:TAGTGTGTATTGGTTTCTCC INRA063 F: ATTTGCACAAGCTAAATCTAACC 5`HEX 52 R:AAACCACAGAAATGCTTGGAAG OarFCB011 F: GCAAGCAGGTTCTTTACCACTAGCACC 5`6-FAM 54 R:GGCCTGAACTCACAAGTTGATATATCTATCAC ILSTS030 F: CTGCAGTTCTGCATATGTGG 5`HEX 54 R:CTTAGACAACAGGGGTTTGG SR-CRSP-1 F: TGCAAGAAGTTTTTCCAGAGC 5`HEX 54 R:GTACCCTGGTTTCACAAAAGG SR-CRSP-5 F: GGACTCTACCAACTGAGCTACAAG 5`6-FAM 54 R:TTATGAAATGAAGCTAAAGCAATGC ILSTS033 F: TATTAGAGGATGGATGCTAAACT 5`HEX 52 R:CTGCAAATAAGAAAACTGAATAAA OarFCB48 F: GAGTTAGTACAAGGATGACAAGAGGCAC 5`HEX 54 R:GACTCTAGAGGTACGCAAAGAACCAG McM038 F: TGGTGAATGGTGCTCTCATACCAG 5`HEX 54 R:CAGCCAGCAGCCTCTAAAGGAC -
使用GeneMarker V2.2.0(Demo)软件,根据DNA片段大小范围和峰形质量确认等位基因。3个重复试验分型一致的结果直接录入数据库,若结果有差异,则需要重新跑PCR甚至重新提取DNA。利用 Excel 2007软件整理所有样品对应微卫星位点的等位基因型数据。
-
利用Micro-Checker[22]检验每个位点是否存在无效等位基因或等位基因缺失等情况。利用Genepop on the Web[23]和GenAlEx[24]软件,对每个微卫星位点是否偏离哈迪-温伯格平衡(Hardy-Weinberg equilibrium,HWE)和是否存在连锁不平衡(Linkage disequilibrium,LD)进行分析。哈迪-温伯格平衡检测中,若P>0.05,则符合哈代-温伯格平衡,说明这个群体是随机交配群体;若P<0.05,则呈显著偏离哈代-温伯格平衡状态;若P<0.01,则呈极显著偏离哈代-温伯格平衡状态。利用Cervus、GenAlEx、Excel Microsatellite Toolkit统计各个微卫星标记位点的等位基因数(k)、有效等位基因数(Ne)、观测杂合度(Observed heterozygosity,Ho)、期望杂合度(Expected heterozygosity,He)和多态信息含量(Polymorphism information content,PIC)[24, 25]。当PIC<0.25、0.25<PIC<0.5和PIC>0.5时,分别表示该座位为低度、中度和高度多态位点,其中高度多态位点能够提供大量的遗传信息,中度多态位点可以提供较为合理的信息[10, 11, 13, 20, 21, 26]。
-
本研究筛选得到的9对多态性微卫星位点中,高度多态标记位点有5对(PIC>0.5),可提供大量的遗传信息,中度多态标记位点有3对(0.25<PIC<0.5),能够提供较为合理的信息,低度多态标记位点仅1对(PIC<0.25),为SR-CRSP-1,可提供的信息量较差(见表2)。9个微卫星标记位点的等位基因数范围是3~7,平均值是4.89;其中,等位基因数是4及以上的微卫星位点有7个,占多态性微卫星位点总数的77.8%;等位基因数在5~7之间的微卫星位点有6个,占多态性微卫星位点总数的66.7%;仅有2个微卫星位点的等位基因数是3个(见表2)。观察杂合度的变异范围是0.250~0.839,平均值是0.547;期望杂合度的变异范围是0.226~0.766,平均值是0.539(见表2)。多态信息含量的变异范围是0.226~0.766,平均值是0.515(见表2)。
依据哈代-温伯格平衡检测中的P值,9个微卫星标记位点中有7个标记位点符合哈代-温伯格平衡,占77.8%;另外2个标记位点(INRA063和SR-CRSP-5)显著偏离哈代-温伯格平衡,占22.2%。
依据连锁不平衡检测结果,9个微卫星标记位点之间均不存在连锁不平衡现象(P>0.001),表明所选用的微卫星标记位点是相互独立的遗传标记系统。
-
排除SR-CRSP-1外之后,使用剩余的8个微卫星标记位点对圈养成都麻羊种群进行遗传多样性分析,共有60个等位基因被成功检测,各标记位点的等位基因数介于5~11之间,平均值为7.5;各标记位点的平均有效等位基因数为2.36个;各个位点私有等位基因检测并未发现其存在;观察杂合度的变异范围是0.413~0.770,平均值是0.577;期望杂合度的变异范围是0.493~0.777,平均值是0.634;多态信息含量的变异范围是0.432~0.743,平均值是0.580(见表3)。
表 2 9个微卫星标记位点多态性检测结果
Table 2. Polymorphism detection results of 9 microsatellite markers
微卫星位点 k N Ho He PIC F(Null) ILSTS004 5 31 0.452 0.435 0.404 −0.0615 INRA063 5 31 0.516 0.654 0.598 0.1032 OarFCB011 4 32 0.594 0.629 0.551 −0.0297 ILSTS030 6 32 0.813 0.685 0.633 −0.0991 SR-CRSP-1 3 32 0.250 0.226 0.226 −0.0608 SR-CRSP-5 7 32 0.594 0.591 0.591 −0.0116 ILSTS033 5 29 0.414 0.459 0.459 0.0122 OarFCB48 6 31 0.839 0.766 0.766 −0.0582 McM038 3 31 0.452 0.410 0.410 −0.0721 注:k表示等位基因数,N表示DNA样品量,Ho表示观察杂合度,He表示期望杂合度,PIC表示多态信息含量 表 3 圈养成都麻羊种群遗传多样性情况
Table 3. Genetic diversity of captive Chengdu gray goat population
微卫星标记 k N Ho He PIC HWE F(Null) ILSTS004 11 260 0.565 0.610 0.585 NS 0.0367 INRA063 5 254 0.594 0.675 0.622 NS 0.0579 OarFCB011 8 260 0.531 0.635 0.565 NS 0.0900 ILSTS030 7 267 0.704 0.696 0.656 NS −0.0110 SR-CRSP-5 8 261 0.598 0.614 0.567 NS 0.0063 ILSTS033 7 262 0.443 0.491 0.468 NS 0.0500 OarFCB48 9 256 0.770 0.777 0.743 NS 0.0037 McM038 5 264 0.413 0.493 0.432 NS 0.0815 注:k表示等位基因数,N表示DNA样品量,Ho表示观察杂合度,He表示期望杂合度,PIC表示多态信息含量,HWE表示哈代-温伯格平衡 -
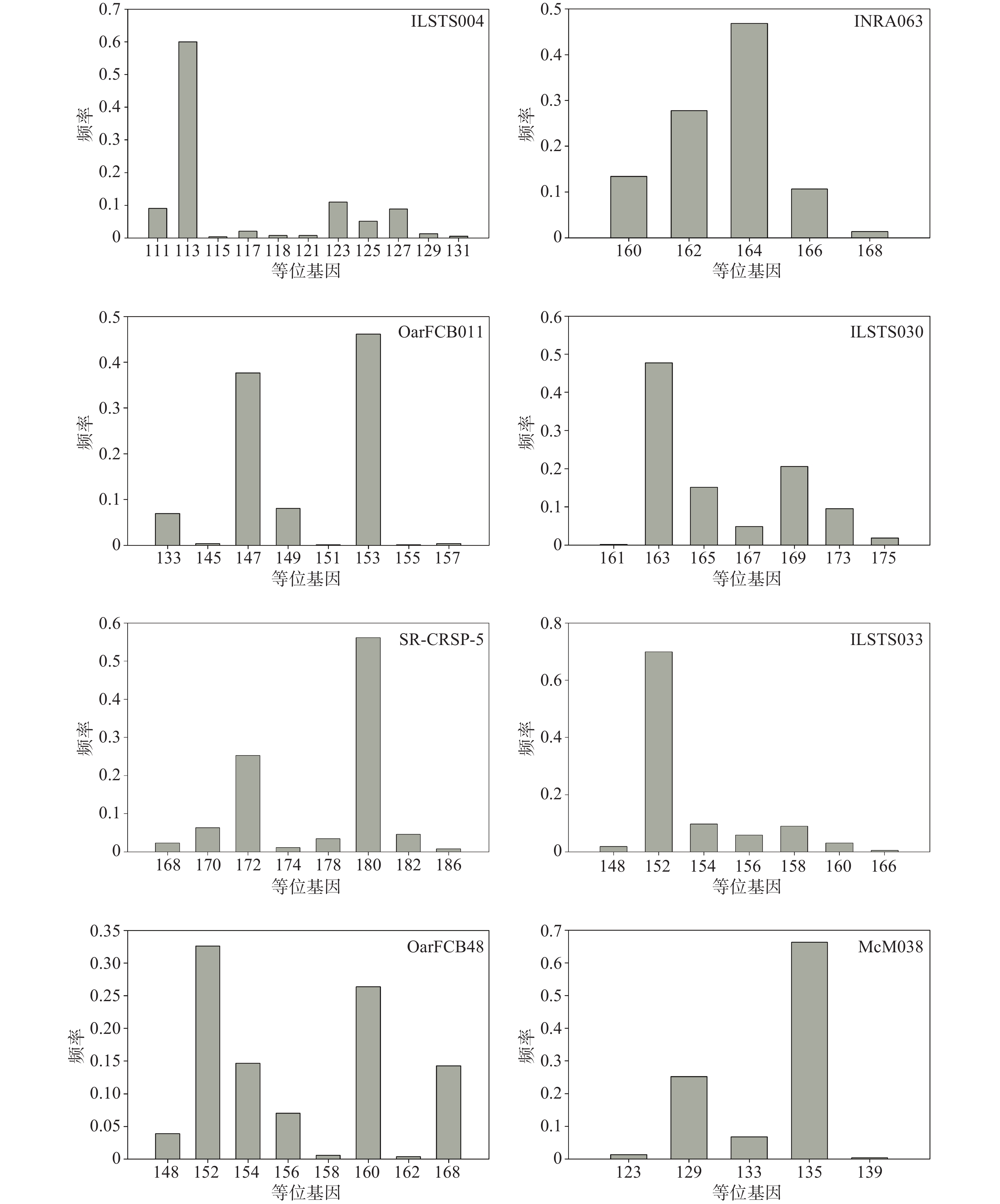
在8个微卫星标记位点中,等位基因较多的前4个是:ILSTS004,共检测到11个等位基因,大小分布在111~131 bp,频率范围为0.0038~0.6000;OarFCB48,共检测到9个等位基因,大小分布在148~168 bp,频率范围为0.0039~0.3262;SR-CRSP-5,共检测到8个等位基因,大小分布在168~186 bp,频率范围为0.0077~0.5613;OarFCB011,共检测到8个等位基因,大小分布在133~157 bp,频率范围为0.0019~0.4615(见图1)。
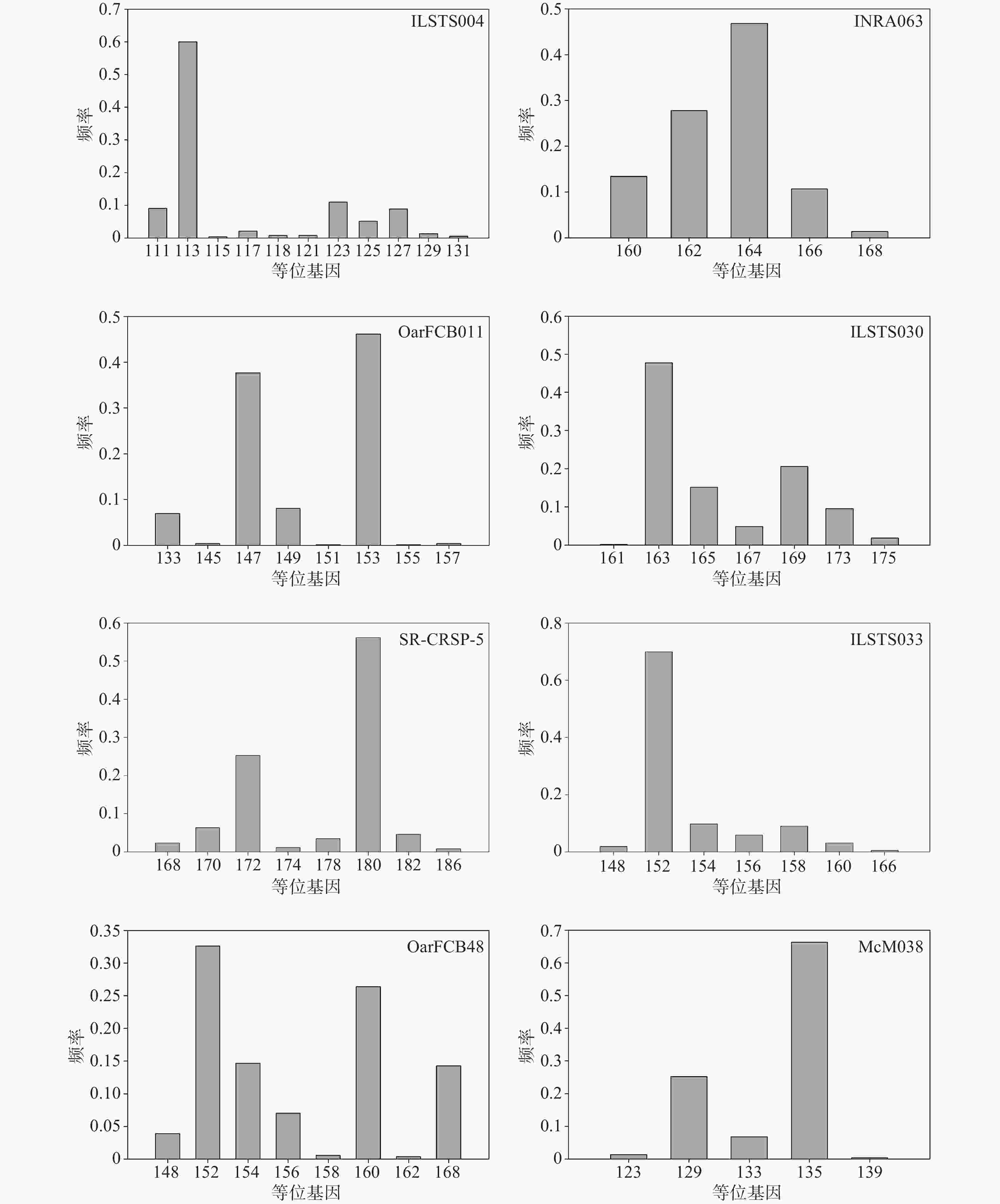
图 1 微卫星标记位点的等位基因分布
Figure 1. Allele frequency of microsatellite markers
在8个微卫星标记位点中,等位基因较少的4个位点是:ILSTS033,共检测到7个等位基因,大小分布在148~166 bp,频率范围为0.0057~0.6985;ILSTS030,共检测到7个等位基因,大小分布在161~175 bp,频率范围为0.0019~0.4775;McM038,共检测到5个等位基因,大小分布在123~139 bp,频率范围为0.0038~0.6629;INRA063,共检测到5个等位基因,大小分布在160~168 bp,频率范围为0.0138~0.4685(见图1)。
-
遗传多样性是一个物种适应环境变化所必需的条件[7]。种群内的遗传多样性反映了一个物种的进化潜力,是其遗传特征进化的基础[26]。一个物种的遗传变异越丰富,其对自然选择和环境变化的适应能力就越强[6, 9, 12, 27]。遗传多样性通常用等位基因多样性、多态性、平均杂合度等指标来描述[6, 12, 16, 19]。
在同一微卫星位点中,随着核心重复序列重复次数的变化,该位点的片段长度也会发生变化,继而形成等位基因[11, 17];等位基因是形成群体遗传多态性的基础,等位基因数越多,遗传多态性越丰富[17]。评价位点多态性高低的指标是多态信息含量[17]。本研究从9个微卫星位点中筛选到5个高度多态的位点、3个中度多态性的位点、1个低度多态性的位点,因此仅有前8个可作为有效的分子标记进行群体遗传多样性分析。在后续的分析中我们使用了这8个中高度多态的微卫星位点分析了268份成都麻羊样品,最终在这8个位点中一共检测到的等位基因数为60个,平均等位基因数为7.5,与其他绵羊种群相比,要高于甘肃高山细尾羊、滩羊、藏羊等[7],低于山东省的小尾寒羊、大尾寒羊、山地绵羊、洼地绵羊等[28],与四川省布拖黑绵羊种群基本持平[10],但与其他山羊种群相比,要高于山西黎城大青羊、山西吕梁黑山羊、宁夏中卫山羊、贵州白山羊、云南威信白山羊等[11, 16, 19],低于湖南湘东黑山羊、重庆川东白山羊、云南云岭山羊、河南黄淮山羊等[11]。不过,这并不能排除种群大小和所用微卫星位点数不同而造成的差异。有研究表明,如果样本量太少,会导致一些频率较低的等位基因无法被检测到,进而对试验结果的准确性造成影响[9]。本研究使用了8个微卫星位点分析了268份成都麻羊样品,微卫星位点数量属于平均水平,种群样本量较大,而个别其他研究的样本量最少仅有20余份[11, 17],微卫星位点数量最少仅有5个[7, 17]。以同一微卫星位点而言,种群样本量越大,其观察到的等位基因数往往越多,以微卫星位点ILSTS004、OarFCB011和ILSTS030为例,本研究中观察到的等位基因数目分别为11、8和7,而在刘成建的研究中(涉及20个样本量)仅为5和4[17]。此外,在本项研究中,成都麻羊群体中8个微卫星位点的多态信息含量的变异范围是0.432~0.743,平均多态信息含量0.580,与刘成建的研究结果(0.604)基本一致[17],表明了成都麻羊群体具有较丰富的遗传多样性。
基因杂合度,它反映了群体在几个位点的遗传变异情况,是被检测的遗传标记的多态性的度量参数[10, 17]。若某个群体的基因杂合度大于0.5,则意味着该群体还未受到高强度的选择压力,其遗传多样性比较丰富[10, 17]。在本项研究中,成都麻羊群体的平均观察杂合度和平均期望杂合度分别为0.577和0.634,表明该群体有较丰富的遗传多样性,该结果与前述多态性的分析结果相一致。另外,刘若余等人基于线粒体D-loop序列调查了中国9个山羊品种的遗传多样性,其中成都麻羊的单倍型多样性和核苷酸多样性分别为1.0000±0.0243和0.017956±0.004410,同样表明其有着丰富的遗传多样性[29]。
有效等位基因数也可以反应群体的遗传变异大小,在数值上等于基因纯合度的倒数[9, 16, 17]。若有效等位基因数越接近所检测到的等位基因的绝对数,则说明等位基因在群体中分布越均匀[9, 16, 17, 21]。本研究中,成都麻羊群体平均有效等位基因数(2.36)低于实际观测到的等位基因数(7.5),说明有些等位基因频率相对较高,而另一些相对较低,等位基因在群体中分布得并不均匀,图1也具体地展示了这一现象,究其原因应该是圈养种群频繁人工选择和一定的近亲繁殖造成的。研究同时发现,成都麻羊群体的观察杂合度和期望杂合度之间差异很小,仅为0.057,这说明就目前而言,成都麻羊群体受外来选择压力和近交繁殖等因素的影响还比较小,群体依旧处于遗传平衡的状态
种群遗传学管理的目的是保持种群的遗传多样性,避免近亲繁殖[30]。如果对一个封闭的小种群不采取任何人工干预措施,最终必将造成近亲交配,进而引发近交衰退。鉴于成都麻羊种群的遗传多样性还比较高,受近亲繁殖等因素影响还较小,具有较高的保种潜力,建议开展全面采样,借助微卫星DNA标记技术,完善个体信息档案,构建起具有完整代表性的成都麻羊种群遗传谱系关系。
Genetic Diversity Assessment of Chengdu Gray Goat Based on Microsatellite Markers
More Information-
摘要: 作为优良的地方山羊品种,成都麻羊对我国山羊品种的改良和新品种培育起了重要作用。畜禽遗传资源本质上是基因资源,保护畜禽遗传资源就是保护基因的多样性。为了了解成都麻羊的遗传情况,通过微卫星标记技术,选用8对中高度多态微卫星标记进行成都麻羊圈养种群的遗传多样性评估,共检测到60个等位基因,平均等位基因和平均有效等位基因数分别为7.5、2.36,平均观察杂合度、平均期望杂合度和平均多态信息含量分别为0.577、0.634和0.580。研究结果表明成都麻羊种群遗传多样性较高,具有较高的保种潜力。Abstract: Chengdu gray goat is an excellent local goat breed, which plays an important role in the improvement of goat breeds and the cultivation of new breeds in China. However, due to the lack of scientific breeding methods, some captive populations of Chengdu gray goat lead to the confusion of pedigree and inbreeding, which, to some extent, will lead to the degradation of the captive population. In the long run, the genetic diversity of Chengdu gray goat will gradually lose and some unique important genetic resources will gradually disappear. In this study, the genetic diversity assessment of Chengdu gray goat was conducted using the microsatellite markers. The results were as follows: (1) Eight pairs of microsatellite markers with medium and high polymorphism were successfully developed by cross-species method; (2) A total of 60 alleles were detected, with an average allele of 7.5, an average effective allele number of 2.36, an average observed heterozygosity of 0.577, an average expected heterozygosity of 0.634, and an average polymorphism information content of 0.580. The results showed that the genetic diversity of Chengdu gray goat population was high and it has high conservation potential.
-
Key words:
- Chengdu gray goat;
- Microsatellite;
- Genetic diversity;
- Inbreeding
-
表 1 本研究所用微卫星DNA标记
Tab. 1 Microsatellite DNA markers were used in this study
微卫星位点 引物序列(5′-3′) 5′端修饰 退火温度/℃ ILSTS004 F: CTTAAAATCTGTCTTTCTTCC 5`6-FAM 48 R:TAGTGTGTATTGGTTTCTCC INRA063 F: ATTTGCACAAGCTAAATCTAACC 5`HEX 52 R:AAACCACAGAAATGCTTGGAAG OarFCB011 F: GCAAGCAGGTTCTTTACCACTAGCACC 5`6-FAM 54 R:GGCCTGAACTCACAAGTTGATATATCTATCAC ILSTS030 F: CTGCAGTTCTGCATATGTGG 5`HEX 54 R:CTTAGACAACAGGGGTTTGG SR-CRSP-1 F: TGCAAGAAGTTTTTCCAGAGC 5`HEX 54 R:GTACCCTGGTTTCACAAAAGG SR-CRSP-5 F: GGACTCTACCAACTGAGCTACAAG 5`6-FAM 54 R:TTATGAAATGAAGCTAAAGCAATGC ILSTS033 F: TATTAGAGGATGGATGCTAAACT 5`HEX 52 R:CTGCAAATAAGAAAACTGAATAAA OarFCB48 F: GAGTTAGTACAAGGATGACAAGAGGCAC 5`HEX 54 R:GACTCTAGAGGTACGCAAAGAACCAG McM038 F: TGGTGAATGGTGCTCTCATACCAG 5`HEX 54 R:CAGCCAGCAGCCTCTAAAGGAC  下载: 导出CSV
下载: 导出CSV
表 2 9个微卫星标记位点多态性检测结果
Tab. 2 Polymorphism detection results of 9 microsatellite markers
微卫星位点 k N Ho He PIC F(Null) ILSTS004 5 31 0.452 0.435 0.404 −0.0615 INRA063 5 31 0.516 0.654 0.598 0.1032 OarFCB011 4 32 0.594 0.629 0.551 −0.0297 ILSTS030 6 32 0.813 0.685 0.633 −0.0991 SR-CRSP-1 3 32 0.250 0.226 0.226 −0.0608 SR-CRSP-5 7 32 0.594 0.591 0.591 −0.0116 ILSTS033 5 29 0.414 0.459 0.459 0.0122 OarFCB48 6 31 0.839 0.766 0.766 −0.0582 McM038 3 31 0.452 0.410 0.410 −0.0721 注:k表示等位基因数,N表示DNA样品量,Ho表示观察杂合度,He表示期望杂合度,PIC表示多态信息含量
下载: 导出CSV
表 3 圈养成都麻羊种群遗传多样性情况
Tab. 3 Genetic diversity of captive Chengdu gray goat population
微卫星标记 k N Ho He PIC HWE F(Null) ILSTS004 11 260 0.565 0.610 0.585 NS 0.0367 INRA063 5 254 0.594 0.675 0.622 NS 0.0579 OarFCB011 8 260 0.531 0.635 0.565 NS 0.0900 ILSTS030 7 267 0.704 0.696 0.656 NS −0.0110 SR-CRSP-5 8 261 0.598 0.614 0.567 NS 0.0063 ILSTS033 7 262 0.443 0.491 0.468 NS 0.0500 OarFCB48 9 256 0.770 0.777 0.743 NS 0.0037 McM038 5 264 0.413 0.493 0.432 NS 0.0815 注:k表示等位基因数,N表示DNA样品量,Ho表示观察杂合度,He表示期望杂合度,PIC表示多态信息含量,HWE表示哈代-温伯格平衡
下载: 导出CSV
-
[1] 张显成,徐成钦. 成都麻羊的历史渊源与性状特点[J]. 四川畜牧兽医,2005,32(10):46. [2] 王杰,欧阳熙,王永,等. 成都麻羊的保种与利用[J]. 西南民族大学学报(自然科学版),2008,34(1):82−86. [3] 王杰,金鑫燕,傅昌秀,等. 成都麻羊与四川各地黑山羊品种(群体)mtDNA D-loop序列多态性研究[J]. 西南民族大学学报(自然科学版),2007,33(2):304−308. [4] 何家良,吴永建,王春秀. 成都麻羊的种质资源保护[J]. 四川畜牧兽医,2014,41(11):16, 19. [5] 陈敏敏,郑劲松,龚成,等. 天鹅洲迁地保护江豚种群近亲繁殖状况评估[J]. 动物学杂志,2014,49(3):305−316. [6] 蒋志刚, 马克平. 保护生物学原理[M]. 北京: 科学出版社, 2014. [7] 柴文琼,成述儒,靳建华,等. 5个绵羊群体微卫星多态性分析[J]. 扬州大学学报(农业与生命科学版),2012,33(1):42−46. [8] 王峥峰. 分子生态学与数据分析基础[M]. 北京: 科学出版社, 2016. [9] 张瑞. 岷县黑裘皮羊微卫星多样性与生产性状的关联分析[D]. 兰州: 甘肃农业大学, 2019. [10] 周明亮,陈明华,庞倩,等. 布拖黑绵羊微卫星标记遗传多样性研究[J]. 草学,2018,238(2):74−80. doi: 10.3969/j.issn.2096-3971.2018.02.014 [11] 贾小姣,陈亚乐,王诗佳,等. 9个山羊品种微卫星DNA遗传多样性分析[J]. 安徽农业大学学报,2019,46(5):779−784. [12] 张永德,曾兰,宾石玉,等. 尼罗罗非鱼选育家系的遗传多样性研究[J]. 广西师范大学学报(自然科学版),2014,32(3):94−101. [13] 丁勇,尚金男,徐怡亮,等. 利用微卫星标记分析安徽地方种猪的遗传多样性[J]. 云南农业大学学报(自然科学),2019,34(5):772−778. [14] 王泽亮,刘青,邢文曦,等. 基于SSR的8个平欧杂交榛品种指纹图谱构建[J]. 四川林业科技,2019,40(2):5−8. [15] 王亚磊,李静心,茆达干,等. 6个绵羊品种微卫星多样性分析[J]. 中国畜牧兽医,2014,41(4):174−179. [16] 张爱玲,马月辉,李宏滨,等. 利用微卫星标记分析6个山羊品种遗传多样性[J]. 农业生物技术学报,2006,14(1):38−44. doi: 10.3969/j.issn.1674-7968.2006.01.008 [17] 刘成建. 四川地方山羊品种遗传多样性及南江黄羊SSRs与生长性状的相关性研究[D]. 雅安: 四川农业大学, 2007. [18] 李晓雨,张纪刚,狄冉,等. 利用微卫星标记分析5个山羊品种的遗传多样性和进化关系[J]. 安徽农业大学学报,2015,42(4):520−528. [19] 朱兰,孙利民,梁家充,等. 利用微卫星标记分析云南省10个山羊品种的遗传多样性[J]. 中国畜牧杂志,2019,55(1):57−63. [20] 张烨华,王立民,王聪慧,等. 6个绵羊品种微卫星多态性分析[J]. 石河子大学学报(自然科学版),2015,33(5):558−562. [21] 张乐超,刘月琴,段春辉,等. 7个地方山羊品种遗传多样性及遗传结构分析[J]. 生物技术通报,2020,36(3):16−23. [22] Van Oosterhout C, Hutchinson W F, Wills D P M, et al. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data[J]. Molecular Ecology Notes, 2004, 4(3): 535−538. [23] Rousset F. genepop’007: a complete re-implementation of the genepop software for Windows and Linux[J]. Molecular Ecology Resources, 2008, 8(1): 103−106. [24] Peakall R, Smouse P E. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research[J]. Molecular Ecology Notes, 2006, 6(1): 288−295. [25] Kalinowski S T, Taper M L, Marshall T C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment[J]. Molecular Ecology, 2007, 16(5): 1099−1106. [26] 孙伟, 常洪. 现代动物群体遗传学[M]. 北京: 科学出版社, 2016. [27] Jangtarwan K, Kamsongkram P, Subpayakom N, et al. Predictive genetic plan for a captive population of the Chinese goral (Naemorhedus griseus) and prescriptive action for ex situ and in situ conservation management in Thailand[J]. PLoS ONE, 2020, 15(6): e0234064. [28] 苑存忠,王建民,马月辉,等. 山东省地方绵羊品种微卫星遗传多态性[J]. 应用生态学报,2006,17(8):1459−1464. doi: 10.3321/j.issn:1001-9332.2006.08.020 [29] Liu R-Y, Yang G-S, Lei C-Z. The genetic diversity of mtDNA D-loop and the origin of Chinese goats[J]. Aata Genetica Sinica, 2006, 33(5): 420−428. [30] 周芸芸,杨万吉,张于光,等. 神农架川金丝猴人工补食群的遗传多样性和亲缘关系[J]. 兽类学报,2015,35(3):229−240. -

 点击查看大图
点击查看大图
图(1) / 表(3)
计量
- 文章访问数: 562
- HTML全文浏览量: 266
- PDF下载量: 41
- 被引次数: 0









