






 下载:
下载:

-
岷江柏(Cupressus chengiana)是柏科柏木属乔木,为国家Ⅱ级重点保护野生植物、川甘特有种,一般生于海拔1200—2 900 m的干燥阳坡[1],其抗逆性强,是干旱河谷地区水土保持和植被恢复的重要乡土树种[2-3]。2014年以来,毛康珊等证实岷江柏依流域分为3个独立单元[4-5],Maerki D.等发表新种甘肃柏木Cupressus gansuensis Maerki & J.Hoch,重新定义大渡河柏木Cupressus fallax Franco,形成“三江三柏”[6],得到国内外植物学界的普遍承认,至此,岷江柏分布区域变窄,仅为汶川、理县、茂县等岷江流域的干旱河谷地区,濒危程度加剧。根据《四川省古树名木目录》(四川省绿化委员会,2020年),现有岷江柏古树12株,分别为阿坝藏族羌族自治州理县6株、茂县6株。经过调查发现,理县现存岷江柏古树仅5株(一级古树3株,二级古树1株,三级古树1株),均得到当地政府妥善管护,生长健康,未发生破坏及砍伐等情况。古树是在气候变化、人为因子、管理措施及传统文化的长期综合作用下保存下来的[7-11],蕴藏有长寿、抗逆、抗病虫害等多种基因的,遗传改良的宝贵种质材料[12-15]。相关研究表明植物的生殖能力一般随着年龄的增长而下降,进而衰老死亡[16],且可能是从花或种子中形成促衰老因子,进而导致其他部位开始衰老[17],在古银杏(Ginkgo biloba)、古楸树(Catalpa bungei)的种子活力及萌发率的研究结果亦是如此[18]。柏科植物具有长寿、生命力旺盛的特点[19],剑阁县古柏的生长状况与树龄无显著相关性[20],古侧柏(Platycladus orientalis)的种子生理机能亦没有明显的衰退迹象[21]。前人已对岷江柏种群结构[22]、种实表型特征[23]、种子萌发[24]、育苗造林[25]、抗性[26-28]、迁地保护[29]等进行了系列研究,而对岷江柏古树的研究鲜见报道,其种子表型、生理特征如何,是否具有繁殖能力尚未可知。因此,为保护古树资源、保存珍贵基因、扩大古树种群数量,以理县岷江柏古树种子为研究对象,通过对种子形态特征、生活力、发芽指数等指标进行测定,研究岷江柏古树种子的萌发特性,探究岷江柏古树种子活力,为古树育苗与珍贵基因保护提供理论基础。
-
2020年9—10月,对研究区域范围内古树(见表1和图1)种子进行采集。选择两年生成熟果,呈绿褐色、鳞片微裂、隐约可见其中褐色种子,利用高枝剪摘果与自然人工承接。采集的球果平铺于阳光照射、通风干燥的房间内晾晒。种鳞开裂后,木棒敲打球果、脱出种子、风扬去杂,去除干瘪、霉变和过小种子。用0.5%的高锰酸钾消毒30 min后用蒸馏水洗净、晾干,装入网袋中存放于室内阴凉通风处[30]。
表 1 岷江柏古树种子采集点及古树信息表
Table 1. Seed collection points and ancient tree information of ancient C. chengiana
采集点 树龄/a 级别 经纬度 海拔/m 树高/m 胸径/cm 冠幅/m 古城 1200 一级 31°33′14.4″N,103°28′54.9″E 1480 42 168 9 城区 600 一级 31°26′21″N,103°10′4.2″E 1860 26 120 11 独柏树 550 一级 31°24′55.8″N 103°7′22″E 1 930 24 71 7 新店子 350 二级 31°26′0.5″N,103°1′46.7″E 2120 32 60 7 
图 1 岷江柏古树
Figure 1. Ancient C. chengiana trees
-
每株古树随机选取100粒种子,用游标卡尺测量种子的长度、宽度及厚度,用百分之一天平测量种子百粒重。每株3个重复,共1200粒种子,并以20树龄岷江柏300粒健康种子为对照,采集地点为四川省阿坝藏族羌族自治州金川县。
-
每株古树挑选外形饱满的种子30粒,参考《1996国际种子检验规程》,开展种子生活力测试。测试方法为氯化三苯基四氮唑(TTC)法,胚、胚乳均染色或胚乳少部分未染色均视为种子有生活力[31]。每株3个重复,共360粒种子,并以90粒健康种子为对照。
-
2021年1月~2月,在四川省林业科学研究院沙河堡实验中心开展不同温度岷江柏古树种子萌发试验。试验开始前,将种子用45 ℃的温水浸泡24 h后,放在铺有2层滤纸的培养皿中分别置于15、20、25 ℃的恒温培养箱(型号:GZX-250光照培养箱)中进行萌发实验。每个温度每株古树选取100粒种子,每个处理3个重复,共1200粒,并以300粒健康种子为对照。
-
当种子胚根长至1 mm时视为发芽,每天10:00观察并记录发芽种子数、始发芽时间,并将发芽种子挑出,计算发芽率、发芽势及发芽指数。试验时间共40 d[25]。
发芽率(%)=(正常发芽的种子粒数/参试种子的总粒数)×100%
发芽势(%)=(发芽试验规定期限的最初1/3期间内的种子发芽数/参试种子的总粒数)×100%
发芽指数=每日发芽数/发芽天数之和
采用单因素方差分析(One-way ANOVA)和LSD(Least significant difference)检验温度处理对古树种子形态特征及萌发的差异性影响[5],绘图采用Excel 2019,指标参数以“平均值±标准误”表示。
-
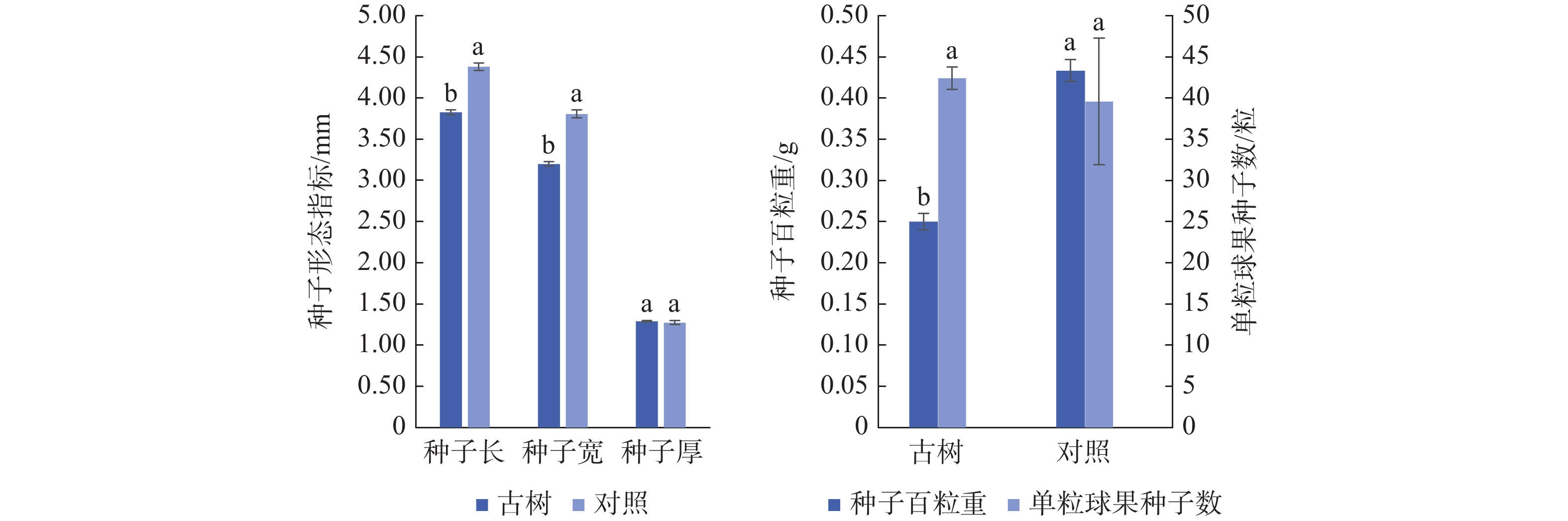
由图2可知,岷江柏古树种子平均长3.83 mm,宽3.20 mm,厚1.29 mm,百粒重0.25 g,单粒球果中平均种子数42.42粒。古树种子长、宽、百粒重均显著(P<0.05)低于对照,而种子厚及单粒球果种子数与对照无显著差异。综合来看,与对照相比,岷江柏古树种子小且质量轻。
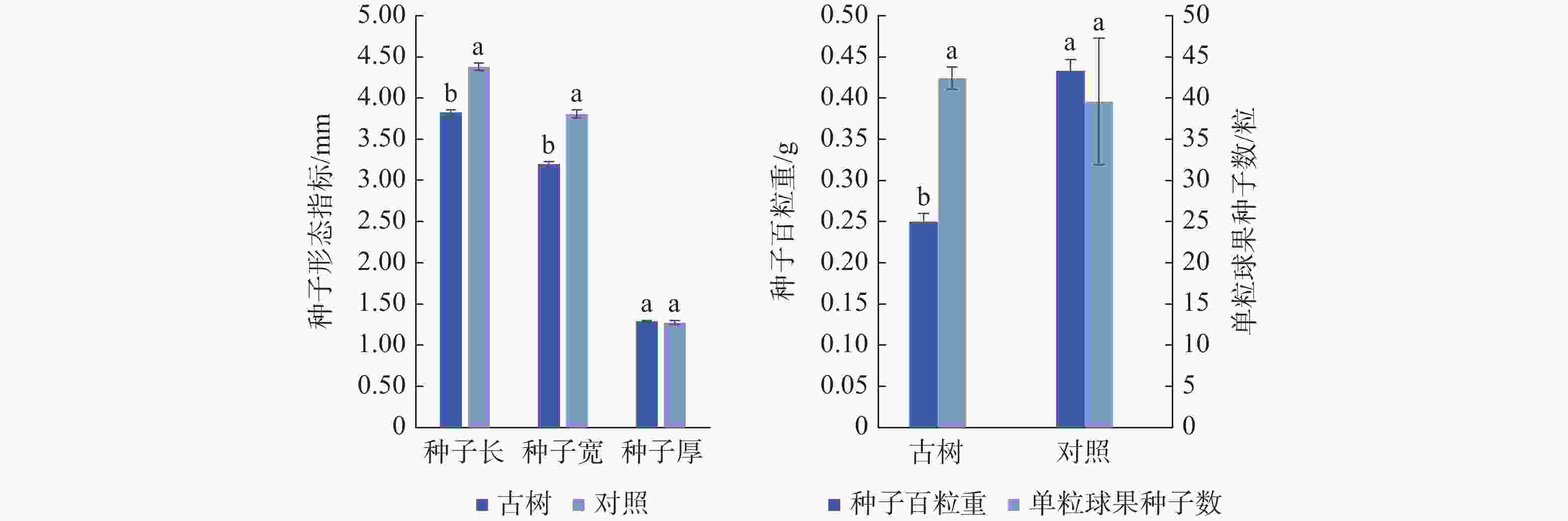
图 2 岷江柏古树种子形态特征
Figure 2. Seed morphological characteristics of ancient C. chengiana trees
-
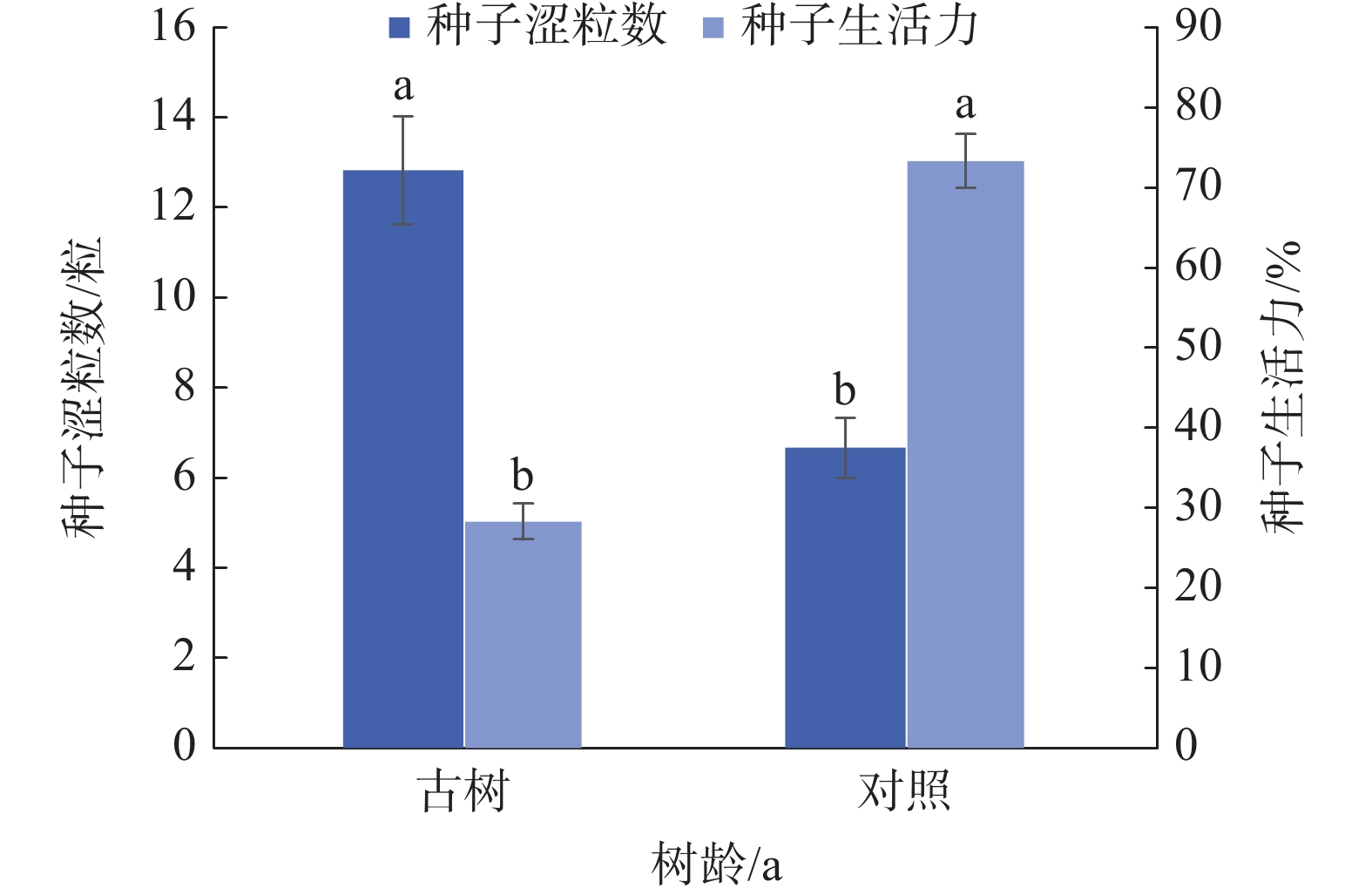
由图3可知,岷江柏古树种子平均涩粒数为12.83粒,显著(P<0.05)高于对照(6.67粒);生活力仅为28.27%,显著(P<0.05)低于对照(73.33%)。综合来看,与对照相比,岷江柏古树种子生活力显著降低。
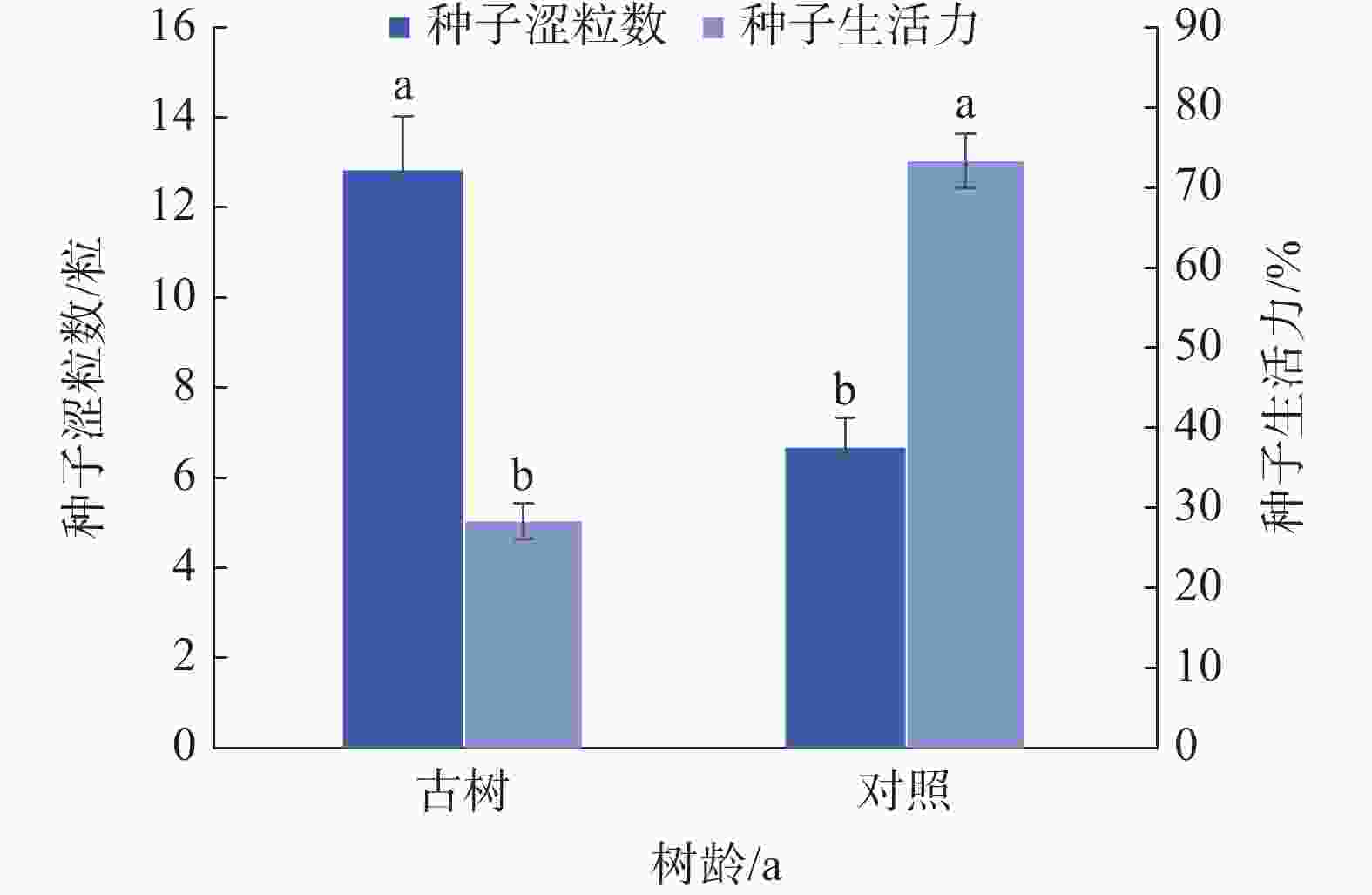
图 3 岷江柏古树种子生活力
Figure 3. Seed viability of ancient C. chengiana trees
-
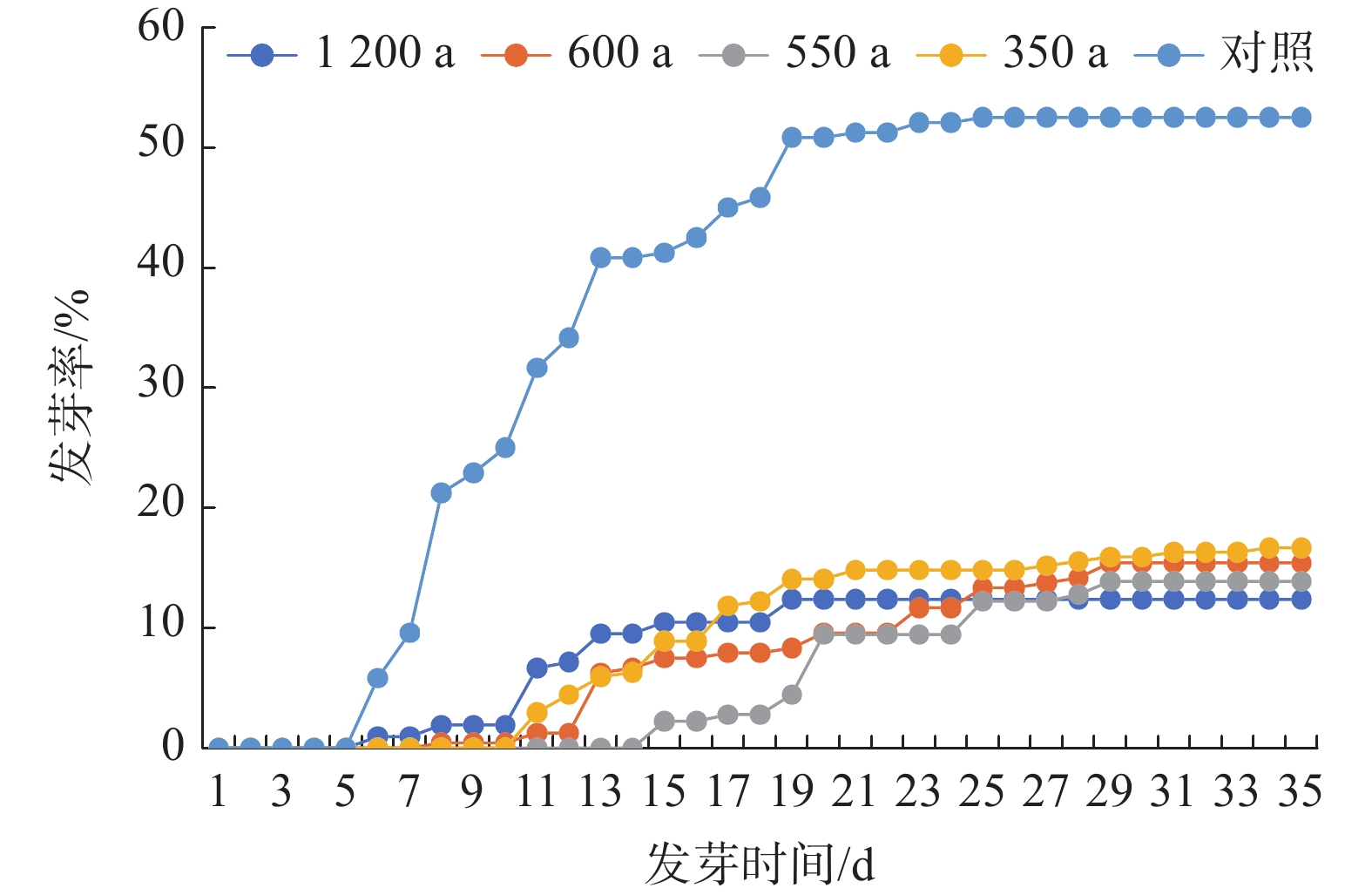
种子发芽率是衡量种子质量好坏的重要指标。由图4可知,岷江柏古树种子平均发芽率为14.59%,始发芽时间为13.75 d,在35 d内完成萌发。根据《林木种子质量分级》(GB 7908—1999)标准,岷江柏古树种子均未达到Ⅲ级种子标准,发芽率低于20%。而对照发芽率为52.50%,平均始发芽时间7 d,并在23 d内即完成萌发,为Ⅰ级种子。综合来看,与对照相比,岷江柏古树种子发芽率低、始发芽时间及发芽周期长。

图 4 岷江柏古树种子萌发特征
Figure 4. Seed germination characteristics of ancient C. chengiana trees
-
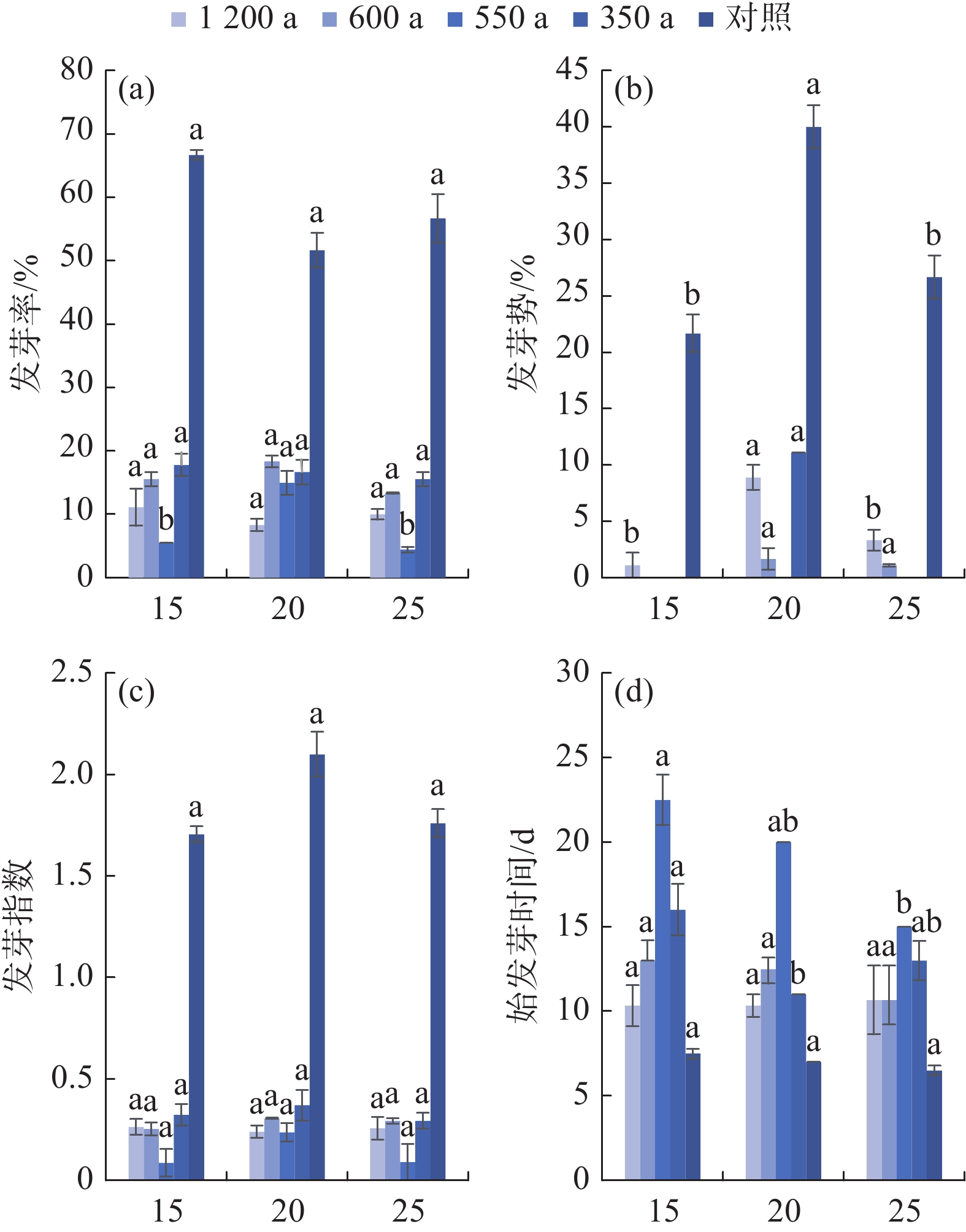
由图5(a)可知,20 ℃下550 a古树种子发芽率显著(P<0.05)高于其他温度处理,1 200 a、600 a、350 a古树种子在不同温度下的发芽率无显著差异。古树种子发芽率均显著(P<0.05)低于对照(58.34%),但古树间发芽率无显著差异(见图5)。20 ℃下,古树种子发芽率最高,平均发芽率为14.58%。350 a古树种子在各温度下发芽情况较好,其次为600 a、1 200 a古树。
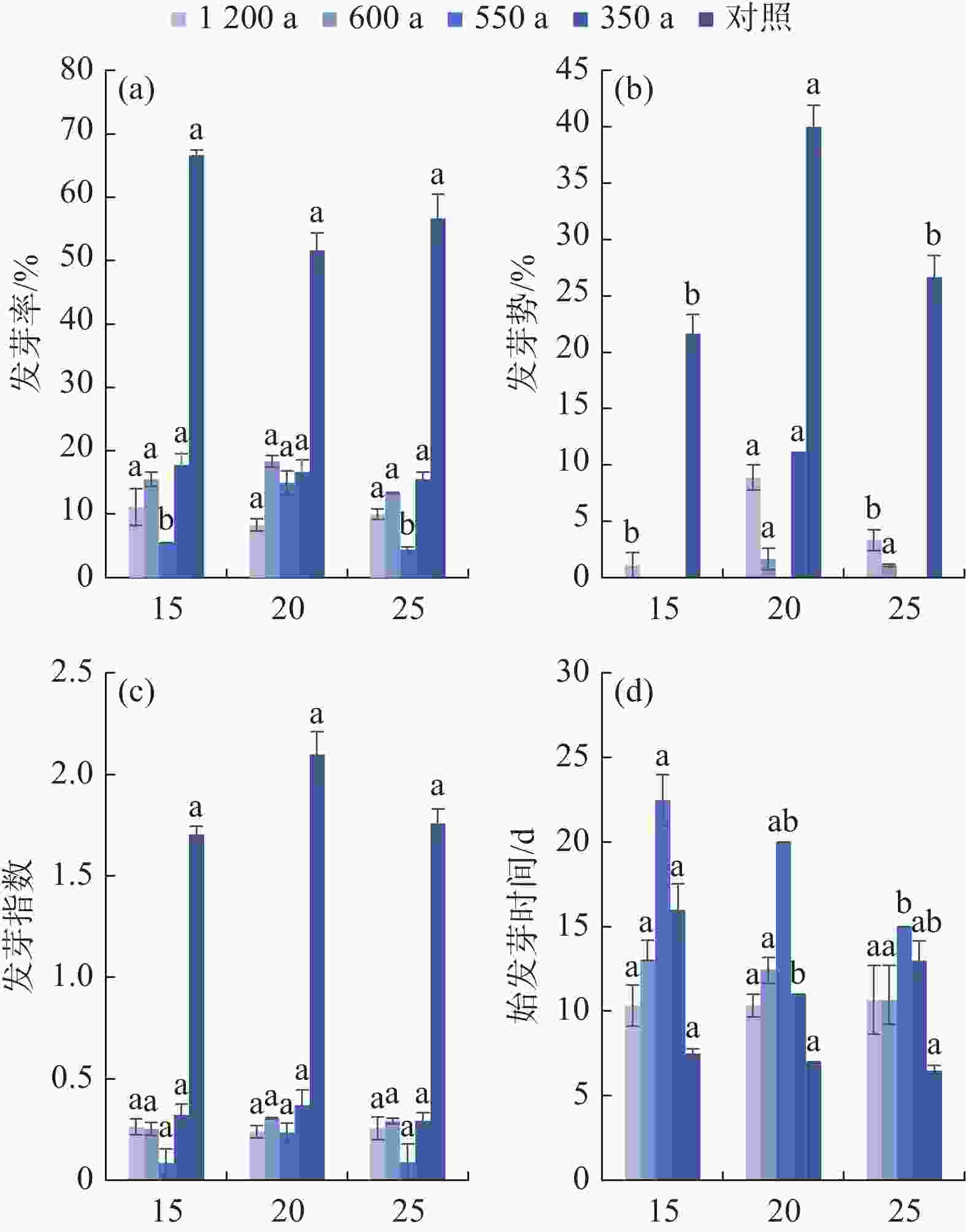
图 5 温度对岷江柏古树种子发芽的影响
Figure 5. Effect of temperature on seed germination of ancient C. chengiana trees
由图5(b)可知,20 ℃下1200 a、350 a古树种子发芽势显著(P<0.05)高于其他温度处理,600 a古树种子发芽势在各温度下差异不显著,550 a古树种子在前10天均未发芽。古树种子发芽势均显著(P<0.05)低于对照(29.45%),但古树间发芽势无显著差异(见图5)。综合来看,古树种子发芽高峰较晚,主要集中在试验的10 d~15 d,而对照在试验第8 d即进入发芽高峰。20 ℃下,古树种子发芽势最高,平均发芽势5.42%;1 200 a古树种子发芽相对更快、更整齐。
由图5(c)可知,古树种子在不同温度下、不同古树间的发芽指数均无显著差异,且均显著(P<0.05)低于对照(1.74)(图5)。综合来看,20 ℃下,古树种子发芽指数最佳,平均发芽指数0.29。350 a古树发芽情况最佳,其次为600 a、1 200 a古树。
由图5(d)可知,25 ℃下550 a古树种子始发芽时间显著(P<0.05)缩短,20℃下350a古树种子始发芽时间显著(P<0.05)缩短,1 200 a、600 a古树种子在不同温度下的始发芽时间无显著差异。古树种子始发芽时间均显著(P<0.05)长于对照(7 d),各古树始发芽时间差异显著(P<0.05),由短及长为1 200 a<600 a<350 a<55 0 a(图5)。综合来看,25℃下,古树种子始发芽时间最短,平均为13.46 d;1 200 a古树种子平均始发芽时间最短,为10.44 d,而550 a古树最长,为19.17 d。
-
岷江柏古树树龄在350—1200年生,均为二级及以上古树,生长在寺庙旁、居民区和公路旁,立地条件差加之人为干扰严重[32],树势差且存在树皮开裂、部分断枝的现象。岷江柏古树种子平均长3.83 mm、宽3.20 mm、厚1.29 mm,百粒重0.25 g,与对照(种子百粒重0.33g)差异显著;单粒球果中平均种子数42.42粒,比对照略有增加(见图2和图3)。可能随着树龄的增加,古树受到的内、外环境胁迫逐渐增加[33],表现出明显的生理功能衰退[16,34],其生长势及繁育能力逐渐下降[35-36],因而结实量少且种子质量轻,存在大量涩粒和空粒。
岷江柏古树种子生活力仅为28.27%,平均发芽率14.59%,与对照(生活力73.33%,发芽率为52.50%)相比,显著降低;始发芽时间(13.75 d)及发芽周期(35 d)显著增加(对照7 d,23 d),未达到Ⅲ级种子标准(见图4)。可能因为古树种子质量轻,代谢活力降低,小种子内部储藏的较少的营养物质使其具有较低的发芽能力和幼苗存活率[17,37]。
不同温度下,古树发芽情况亦无显著差异;1200年生古树始发芽时间显著低于其他古树,不同古树间发芽率、发芽势、发芽指数无显著差异(见图5)。总体来看,岷江柏古树种子的较适宜发芽温度为20 ℃,种子发芽率、发芽势、发芽指数分别为14.59%、5.42%、0.29(对照51.67%、40.00%、2.10),平均始发芽时间13.46 d(对照7 d)。种子萌发受母树发育、种子大小、环境条件等多方面影响[38],岷江柏古树的适宜发芽温度与理县4月的平均气温相近,是植物对生境长期适应的结果[25,39-40]。在酶促反应过程和呼吸作用中,适宜的温度使相应的酶具有较高的活性,种子内部储存的营养物能迅速成为胚能利用的可溶状态,从而促进种子快速萌发[41]。温度过低,酶活性降低,物质转化受阻,发芽率显著降低;温度过高,虽然种子始发芽时间显著缩短但芽弱,高温度导致呼吸高消耗,从而导致有毒物质累积[42],种子萌发受阻。350 a古树树龄较小,种子活力高,发芽率及发芽指数较高;而1200 a古树可能是因为生长海拔较低,植株生长季较长,种子所需的营养生长和发育时间增加[38],种子百粒重大,因而发芽势较高,始发芽时间较短。树龄是影响种子萌发能力的主要因素[21],但因样本数有限,不足以详细诠释树龄对种子萌发的影响,部分内容将在未来的研究中完善。
古树的衰老是树体特征、内部结构及生理功能衰退的综合表现,岷江柏古树结实量少、种子质量轻及萌发能力显著下降,说明岷江柏古树的繁育能力有一定程度的下降,但古树是否已经进入衰老,还需从叶片结构、光合能力、内源激素等方面综合分析。
Seed Morphology and Germination Characteristics of Rare and Endangered Plant Cupressus chengiana
More Information-
摘要: 岷江柏(Cupressus chengiana)是中国珍稀濒危种、国家Ⅱ级重点保护野生植物、川甘特有种。岷江柏古树蕴藏多种基因,是宝贵的遗传种质材料。以阿坝州理县4株岷江柏古树(350—1200年生)种子为试材,以金川岷江柏(20年生)种子为对照,通过研究岷江柏古树种子形态特征及其萌发特性,揭示其种子的萌发特性。结果表明:岷江柏古树平均种子长3.83 mm,宽3.20 mm,厚1.29 mm,单粒球果中平均种子数42.42粒。与对照相比,种子百粒重、生活力、发芽率均显著下降,仅为0.25 g、28.27%、14.59%;始发芽时间、发芽周期(13.75 d、35 d)显著增加,表现出明显的生理功能衰退。1200年生古树始发芽时间显著低于其他古树,不同古树间发芽率、发芽势、发芽指数无显著差异。古树种子较适宜的发芽温度为20℃,发芽率、发芽势、发芽指数分别为14.59%、5.42%、0.29,平均始发芽时间13.46 d。Abstract: Cupressus chengiana is a rare and endangered species in China, a national class II key protected wild plant and endemic to Sichuan and Gansu Province. The ancient C. chengiana trees contain a variety of genes and are valuable genetic germplasm materials. The seeds of 4 ancient C. chengiana trees (350-1200 years) in Lixian County, Aba Prefecture were used as test materials, and the seeds of C. chengiana (20 years old) in Jinchuan County was used as control. Based on the study of the seed morphological characteristics and germination characteristics of ancient C. chengiana, the germination characteristics of their seeds were revealed. The results showed that the average seed length, width and of thickness of ancient C. chengiana trees was 3.83mm, 3.20mm and 1.29mm, and the average number of seeds in a single cone was 42.42. Compared with the control, the 100-seed weight, viability and germination rate of seeds decreased significantly, which were only 0.25g, 28.27%, 14.59%. The initial germination time and the germination period (13.75 days and 35 days) increased significantly, showing obvious physiological function decline. The initial germination time of 1200-year-old ancient trees was significantly lower than that of other ancient trees, and there was no significant difference in germination rate, germination potential and germination index among different ancient trees. The suitable germination temperature of ancient tree seeds was 20 ℃, the germination rate, germination potential and germination index were 14.58%, 5.42% and 0.29 respectively, and the average initial germination time was 13.46 days.
-
图 2 岷江柏古树种子形态特征
注:不同小写字母表示差异显著,P<0.05。
Fig. 2 Seed morphological characteristics of ancient C. chengiana trees
图 5 温度对岷江柏古树种子发芽的影响
Fig. 5 Effect of temperature on seed germination of ancient C. chengiana trees
表 1 岷江柏古树种子采集点及古树信息表
Tab. 1 Seed collection points and ancient tree information of ancient C. chengiana
采集点 树龄/a 级别 经纬度 海拔/m 树高/m 胸径/cm 冠幅/m 古城 1200 一级 31°33′14.4″N,103°28′54.9″E 1480 42 168 9 城区 600 一级 31°26′21″N,103°10′4.2″E 1860 26 120 11 独柏树 550 一级 31°24′55.8″N 103°7′22″E 1 930 24 71 7 新店子 350 二级 31°26′0.5″N,103°1′46.7″E 2120 32 60 7  下载: 导出CSV
下载: 导出CSV
-
[1] 四川森林编辑委员会. 四川森林[M]. 北京:中国森林出版社,1990. [2] 范建容,杨超,包维楷,等. 西南地区干旱河谷分布范围及分区统计分析[J]. 山地学报,2020, 38(2):303−313. [3] 王卫霞,罗达,史作民,等. 岷江干旱河谷造林对土壤微生物群落结构的影响[J]. 生态学报,2014, 34(4):890−898. [4] Xu T T, Wang Q, Olson M S, et al. Allopatric divergence, demographic history, and conservation implications of an endangered conifer Cupressus chengianain the eastern Qinghai-Tibet Plateau. Tree Genetics & Genomes, 2017. [5] Li J, Milne R I, Ru D, et al. Allopatric divergence and hybridization within Cupressus chengiana (Cupressaceae), a threatened conifer in the northern Hengduan Mountains of western China[J]. Molecular Ecology, 2020, 29(7). [6] Maerki D, Hoch J. Taxonomy of the cypresses of Sichuan and Gansu. Bulletin of the Cupressus Conservation Project, 2020, 9: 3-12. [7] 曲凯,李际红,国浩平,等. 山东省流苏古树资源及其保护对策[J]. 山东农业大学学报(自然科学版),2020, 51(5):818−824. [8] HARTEL T, HANSPACH J, MOGA C I, et al. Abundance of large old trees in wood-pastures of Transylvania (Romania)[J]. Science of the Total Environment, 2018, 613/614: 263−270. doi: 10.1016/j.scitotenv.2017.09.048 [9] LAI P Y, JIM C Y, TANG G D, et al. Spatial differentiation of heritage trees in the rapidly-urbanizing city of Shenzhen, China[J]. Landscape and Urban Planning, 2019, 181: 148−156. doi: 10.1016/j.landurbplan.2018.09.017 [10] LINDENMAYER D B, BLANCHARD W, BLAIR D, et al. Environmental and human drivers influencing large old tree abundance in Australian wet forests[J]. Forest Ecology and Management, 2016, 372: 226−235. doi: 10.1016/j.foreco.2016.04.017 [11] ZHANG H, LAI P Y, JIM C Y. Species diversity and spatial pattern of old and precious trees in Macau[J]. Landscape and Urban Planning, 2017, 162: 56−67. doi: 10.1016/j.landurbplan.2017.02.002 [12] 纵丹,周安佩,张垚,等. 西南地区古杨树遗传多样性的SSR分析[J]. 西北植物学报,2018, 38(5):839−849. [13] LINDENMAYER D B, LAURANCE W F, FRANKIN J F. Global decline in large old trees[J]. Science, 2012, 338(6112): 1305−1306. doi: 10.1126/science.1231070 [14] FAISON E K. Large old tree declines at broad scales: a more complicated story[J]. Conservation Letters, 2014, 7(1): 70−71. doi: 10.1111/conl.12075 [15] 詹运洲,周凌. 生态文明背景下城市古树名木保护规划方法及实施机制的思考——以上海的实践为例[J]. 城市规划学刊,2016(1):106−115. [16] STEWART E J, MADDEN R, PAUL G, et al. Aging and death in an organism that reproduces by morphologically symmetric division[J]. PloS Biology, 2005, 3(2): 295−300. [17] 常二梅. 侧柏古树抗衰老分子机理研究[D]. 中国林业科学研究院,2012. [18] MUNNE-BOSCH S. Do perennials really senesce[J]. Trends Plant Science, 2008, 13(5): 216−220. doi: 10.1016/j.tplants.2008.02.002 [19] 李永正,赵鹏鹏. 古柏迁地保护移植关键技术要点浅析[J]. 南方农业,2020, 14(27):32−33. [20] 伏建芳,唐天勇,何显平,等. 剑阁县驿道古柏资源调查初报[J]. 四川林业科技,2018, 39(6):97−101. [21] 常二梅,史胜青,刘建锋,等. 古侧柏种子活力与树龄的关系研究[J]. 西北植物学报,2012, 32(1):166−172. [22] 袁志忠. 岷江柏四个地理种群分布格局比较研究[J]. 中国野生植物资源,2009, 28(5):20−23+44. doi: 10.3969/j.issn.1006-9690.2009.05.005 [23] 冯秋红,史作民,徐峥静茹,等. 岷江柏天然种群种实表型变异特征[J]. 应用生态学报,2017, 28(3):748−756. [24] 徐峥静茹,汪清平,冯秋红,等. 温度对不同种源地岷江柏种子发芽的影响[J]. 西部林业科学,2017, 46(2):107−112. [25] 李东胜,罗达,史作民,等. 四川理县杂谷脑干旱河谷岷江柏造林恢复效果评价[J]. 生态学报,2014, 34(9):2338−2346. [26] 李登峰,魏仕军,陈静,等. 岷江上游干旱河谷岷江柏的光合与水分生理特征干湿季对比研究[J]. 生态学报,2022, 42(18):7381−7389. [27] 冯秋红,缪国辉,徐峥静茹,等. 施氮对干旱河谷岷江柏(<italic> Cupressus chengiana</italic>)幼苗光合生理特征的影响[J]. 西南农业报,2020, 33(7):1455−1460. [28] 邵芳丽,宫渊波,关灵,等. 不同水氮条件对岷江柏幼苗生长的影响[J]. 水土保持通报,2012, 32(1):45−49+53. [29] 常二梅,刘建锋,黄跃宁,等. 岷江柏野生居群和迁地保护居群的遗传多样性比较[J]. 植物研究,2022, 42(5):772−779. [30] 秦爱丽,郭泉水,马凡强,等. 温度、光照和水分对珍稀濒危树种崖柏种子萌发的影响[J]. 种子,2020, 39(2):15−20. [31] 国际种子检验协会(ISTA). 1996国际种子检验规程[M]. 北京:中国农业出版社,1999. [32] MUNNÉ, LALUEZA P. Age-related changes in oxidative stress markers and abscisic acid levels in a drought tolerant shrub, <italic> Cistus clusii</italic> grown under Mediterranean field conditions[J]. Planta, 2007, 225(4): 1039−1049. doi: 10.1007/s00425-006-0412-z [33] 张艳洁,丛日晨,赵琦,等. 适用于表征古树衰老的生理指标[J]. 林业科学,2010, 46(3):134−138. [34] Monaghan P, Charmantier A, Nussey D H, et al. The evolutionary ecology of senescence[J]. Functional Ecology, 2008, 22(3): 371−378. doi: 10.1111/j.1365-2435.2008.01418.x [35] LU T, FINKEL T. Free radicals and senescence[J]. Experimental Cell Research, 2008, 314(9): 1918−1922. doi: 10.1016/j.yexcr.2008.01.011 [36] Passos J F,Nelson G,Wang C,et al. Feedba对照 between p21 and reactive oxygen production is necessary for cell senescence[J]. Molecular Systems Biology,2010, 6:347. [37] Larios E, Alberto Birquez, Becerra J X, et al. Natural selection on seed size through the life cycle of a desert annual plant [J]. Ecology, 2014, 95(11). [38] 杨紫玲,喻舞阳,郑维列,等. 薄毛海绵杜鹃结实和萌发特性随环境的变化[J]. 生态学报,2021, 41(1):101−109. [39] 许静,李文龙,吴鑫悦,等. 青藏高原高寒草甸种子萌发行为沿海拔梯度的分异特征[J]. 草地学报,2021, 29(S1):10−18. [40] 崔现亮,罗娅婷,毕廷菊,等. 储藏和萌发温度对青藏高原东缘12种灌木种子萌发的影响[J]. 生态学杂志,2014, 33(1):23−32. [41] 王艳梅,张小雪,朱秀征,等. 林木种子休眠与萌发机制研究进展[J]. 林业科学,2021, 57(10):128−144. [42] 高荣岐,张春庆. 种子生理学[M]. 北京:中国农业出版社,2009. -

 点击查看大图
点击查看大图

图(5) / 表(1)
计量
- 文章访问数: 179
- HTML全文浏览量: 76
- PDF下载量: 36
- 被引次数: 0









