






 下载:
下载:



-
草原生态系统是我国最大的陆地生态系统,具有调节气候、涵养水源、碳固存、生物多样性保育等不可替代的服务功能[1]。我国草原面积占陆地总面积的42%,是重要的畜牧业生产基地和绿色生态屏障,在保障国家生态安全、粮食安全乃至全球生态平衡中发挥重要作用[1]。尽管气候预测模型结果存在较大的不确定性,但联合国政府间气候变化专门委员会(IPCC)第六次气候科学报告《气候变化2021:自然科学基础》明确指出,在全球气候变暖的背景下,干旱、半干旱地区极端干旱事件发生的频率、强度和持续时间将会显著增加[2],对草甸草地生态系统的结构、功能产生了深远影响。尽管极端干旱事件持续时间较短,但其对植物群落和生态系统的影响可能会超越多年持续性普通干旱的影响[3,4]。因此,21世纪以来,极端干旱如何影响草甸草地生态系统以及草地生态系统如何响应极端干旱成为生态学家和植物学家的关注焦点。
极端干旱事件后,土壤水分与养分等关键资源极度下降,物种间竞争增强,植物迅速占据并利用资源[4, 5]。在长期的进化过程中,草原植物对干旱和半干旱的水分资源波动进化出了各自的适应策略,能够通过改变自身的形态和生理机能提高水分利用效率,适应不同的水分状况,提高自身的竞争能力与生态适应性[6]。在面临极端干旱干扰时,不同物种将采取不同适应策略来应对极端干旱事件[7]。有些物种选择将更多的资源分配到地上部分用于防御环境胁迫(逃避策略),而有些物种将更多的资源贮存于根部用于干旱过后的再生长(忍耐策略)。在干旱发生后,由于忍耐策略物种的超强再生恢复力,逐渐排挤逃避策略的物种,从而成为群落的优势种[8]。
植物功能性状组成是与植物获取和利用资源能力息息相关的属性或特征,最大限度地反映植物对环境变化的适应性信息[9]。因此,群落功能多样性在一定程度上反映物种在资源利用上的生态位互补,可用于探索植物群落应对极端干旱事件的潜在生态学机制[10]。群落功能性状组成的变化由物种内和物种间性状变异共同组成,前者表示种群内个体间性状的差别,后者表示物种间性状的差别[5]。理论上认为,群落水平上多维性状结构组成越复杂,植被在多维生态空间中的配置越多元化,资源配置与利用越合理充分,则植物群落结构和功能的稳定性就越强,反之亦然[11]。因此,基于功能性状的视角,探讨干旱事件对草甸草原生态系统结构和功能的影响不仅可以丰富植被生态适应机理的理论研究,而且对于草甸草原应对干旱事件,开展草地保护与合理利用具有重要的指导意义。
本研究以内蒙古草甸草原植物群落为研究对象,通过控制生长季降雨的干旱模拟实验,旨在探究植物群落性状对干旱胁迫的响应及其对生态系统功能的影响,为预测气候变化下草原生态系统结构和功能提供科学依据。
-
研究地点位于内蒙古自治区额尔古纳市境内的中国科学院沈阳应用生态研究所额尔古纳森林草原过渡带生态系统研究站(50°10′46″ N,119°22′56″ E)。该区域为寒温带大陆性季风气候,夏季温暖多雨,冬季寒冷少雨,雨热同期,年均降雨量为362 mm,年均温度为−2.4 °C。该区地带性植被为草甸草原,天然优势物种内有多年生根茎型禾草羊草(Leymus chinensis)、多年生丛生型禾草贝加尔针茅(Stipa baicalensis)、多年生杂类草披针叶黄华(Thermopsis lanceolata)和细叶白头翁(Pulsatilla turczaninovii)。土壤类型为黑钙土,pH值为7.9。
-
在实验区域内选取相对均质(土壤、植被等)并具有典型代表性的地段,搭建遮雨棚(见图1)。实验采取随机区组设计,包括两种实验处理,分别为对照(自然降雨)和极端干旱(减少6—7月生长季降雨量的100%)。每个处理重复6次,共12个小区。实验小区面积为6×6 m,相邻小区间隔2 m。小区四周挖1 m深,用长6 m和宽1 m的塑料阻隔,防止水分侧向移动。样方中心4×4 m区域为实验采样区,周边设有1 m缓冲区,降低边缘效应。实验采用透明丙烯酸塑料板减少实验样地的降雨量。该材料高透光性,低泛黄度指数,高UV穿透,既能达到减少降水的效果,又不会降低光照强度。遮雨棚顶部采用田字形钢架结构,由4个3×3 m的样框组成,共遮挡36 m2的遮雨面积。每个样框以样方边缘为边,向上倾斜15°,在中间形成尖顶,尖顶长度为6 m,顶点高度为2.5 m,两侧遮雨板尾端距地面60 cm(见图1)。该结构设计有利于所截降水的流走,并保障空气流通,排除温室效应。遮雨棚于每年6月1日安装,7月31日拆除。经过连续4年(2015—2018)的实验处理,干旱持续时间和强度均已达到历史极端干旱事件水平。

图 1 草原极端干旱控制实验
Figure 1. Extreme drought manipulation experiment in grassland
-
于每年8月中旬生长季旺期,在每个实验采样区内随机设置1×1 m样方,进行植物群落种类、植被高度调查,齐地面剪下地上植株部分,装入已编号信封中带回室内,烘干后称重。植物叶片样品的采集根据物种调查的结果,收集样方群落调查内出现的所有物种的叶片,测定每个物种的最大高度、比叶面积、干物质含量和叶片碳、氮、磷含量5个功能性状。
植物功能性状测量方法如下:
最大高度:植株顶端与地面的自然垂直距离,用刻度尺进行测量;
比叶面积:叶片面积与干重的比值。采集植物熟个体,剪下置于自封袋后放入纯净水中浸泡,使其充分吸水,进行叶面积测量,而后,65°C烘干称重;
叶干物质含量:叶片干重与叶片饱和鲜重的比值。叶干物质含量与比叶面积的测定同时进行,将扫描完的叶片用万分之一天平称鲜重,然后,65°C烘干称重;
叶片碳氮磷:碳氮采用元素分析仪进行测定,磷采用钼锑抗比色法进行测定。
-
群落功能多样性包括群落加权性状及功能多样性指数(Laliberté and Legendre, 2010),其计算过程均在R软件的“FD”语言包(package)中实现,公式如下:
$$ {\rm{CWM}}=\frac{\displaystyle\sum\nolimits _{i=1}^{s}{p}_{ij}\times {t}_{ij}}{\displaystyle\sum\nolimits _{i=1}^{S}{a}_{ij}} $$ 式中,CWM(Community-weighted mean traits)为群落加权性状值(一维),tij代表物种i在群落j中的性状值,pij代表物种i在群落j中的相对多度,S为群落j中的物种数。
FDis为多维功能离散指数,通过定义利用物种计算的凸多边形体积的重心,计算每一个物种距离重心的平均距离,最后利用相对多度进行权重处理得到功能离散度,公式如下:
$$ {\rm FDis}=\frac{d+\widehat{dG}}{\left|d\right|+\widehat{dG}} $$ 其中,
$$ d={\sum }_{i=1}^{S}pi\times \left(dGi-\widehat{dG}\right) $$ 式中,FDis(functional divergence)指功能离散度(多维),
$ \widehat{dG} $ 是欧氏距离,根据凸多边形方法,即计算各物种性状距离该凸多边形重心点的距离差之平方和开方获得,$ \widehat{dG} $ 是各物种性状欧式距离的均值,$ \left|d\right| $ 是对$ d $ 计算时取距离差绝对值所得,pi为物种i的相对多度。采用混合效应模型方差分析,对实验处理对植物群落性状加权均值和多样性的影响进行检验,此外对植物群落性状加权均值和多样性与地上净初级生产力之间的关系进行回归分析并检验。本文中的数据统计分析均在R(3.6.1)软件中进行。
-
经过连续4年极端干旱处理,植物群落权重性状(高度、比叶面积、干物质含量、叶碳、氮和磷含量)均值都没有发生显著变化(见图2)。同样,极端干旱处理对植物群落多维性状多样性的影响不具有显著性(见图3)。

图 2 极端干旱对植物群落性状权重均值的影响
Figure 2. Effects of extreme drought on plant community-weighted trait means

图 3 极端干旱对植物群落性状多样性的影响
Figure 3. Effects of extreme drought on plant community trait diverity
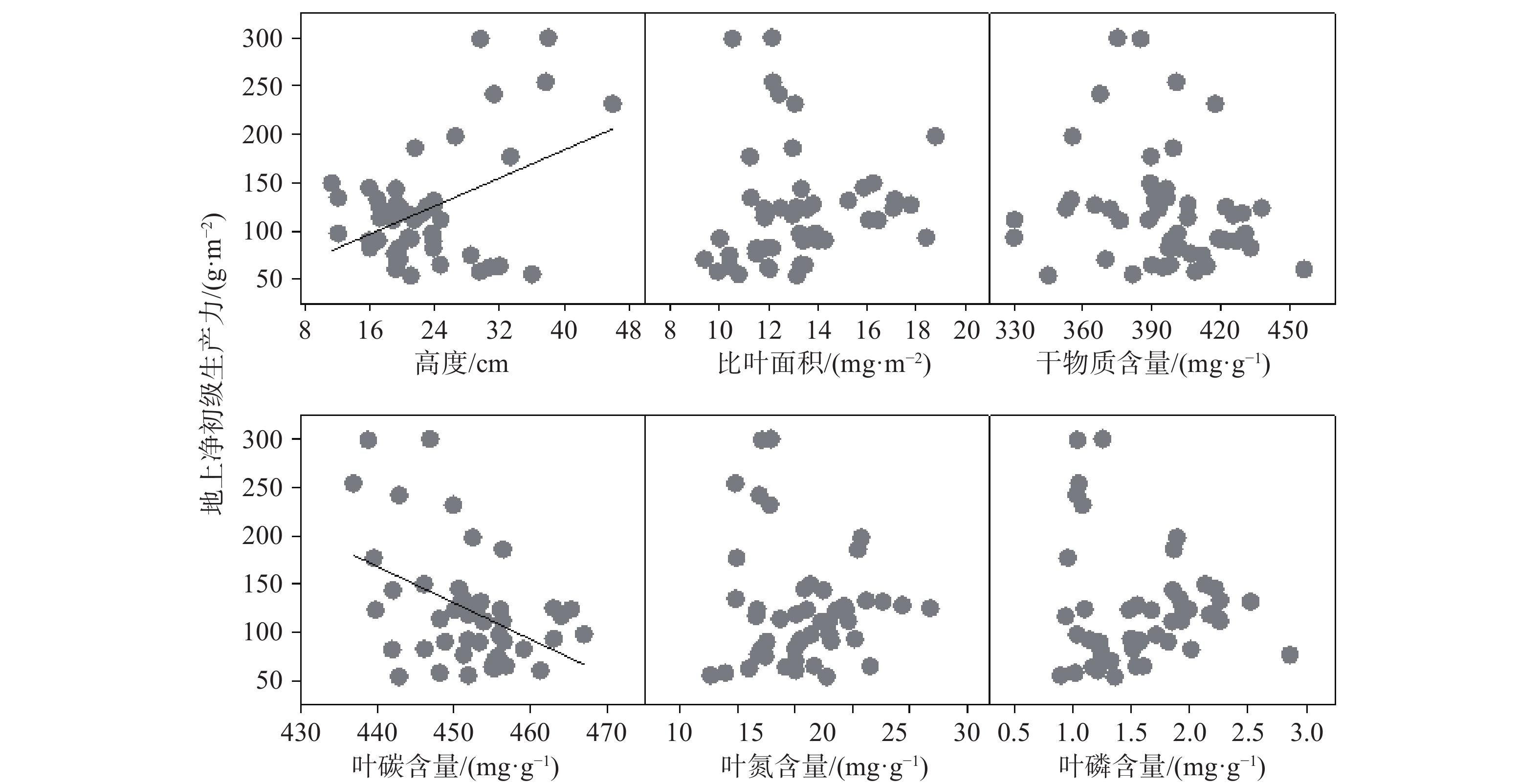
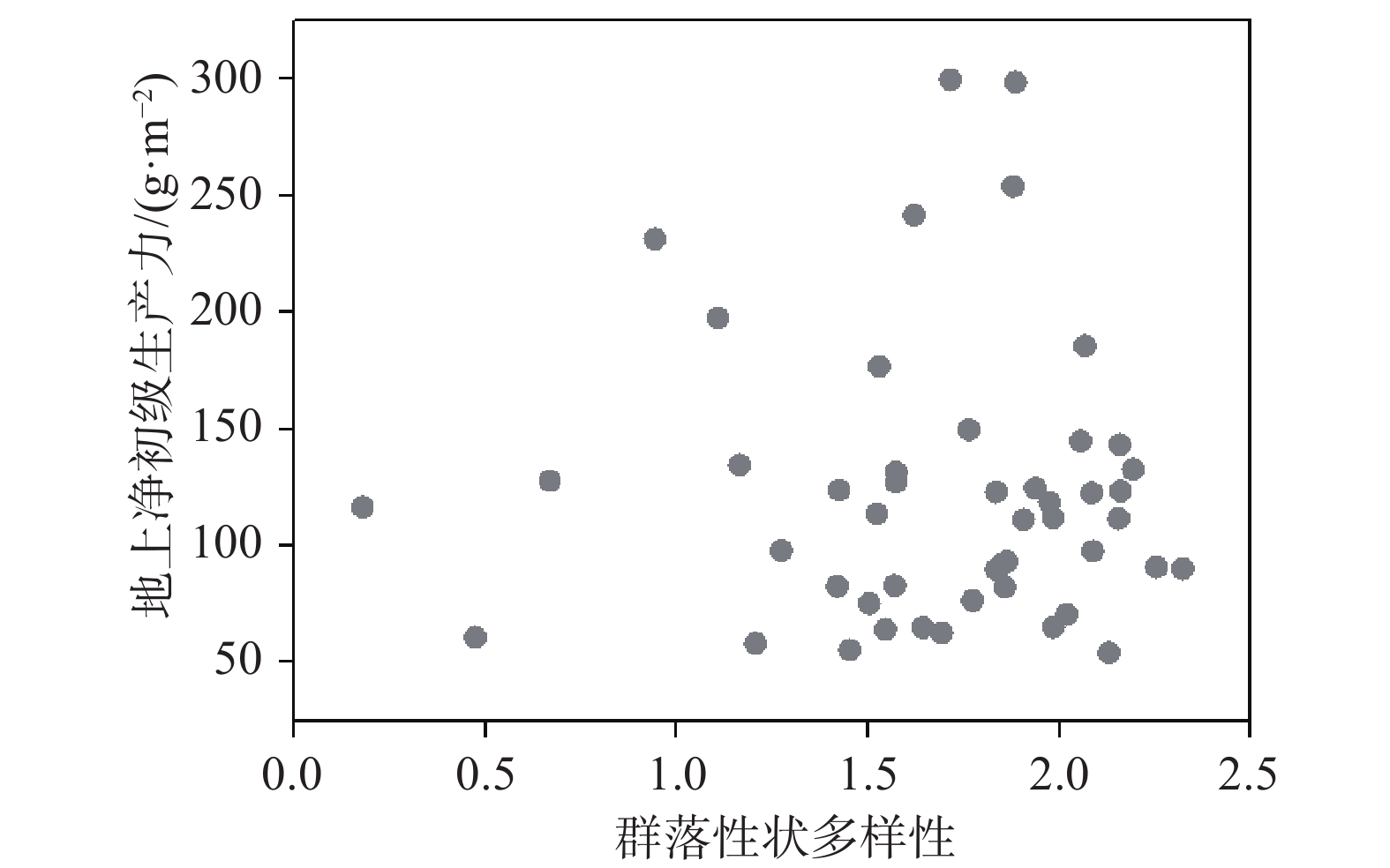
极端干旱条件下,植物群落地上净初级生产力与植物群落权重高度之间呈显著正相关关系,生产力随群落高度的升高逐渐增加。植物群落地上净初级生产力与植物群落权重叶碳含量极端干旱条件下呈显著负相关关系,生产力随群落叶碳含量的增加逐渐降低。极端干旱条件下,植物群落地上净初级生产力与其他经济性状(比叶面积、干物质含量、叶氮和叶磷含量)之间没有显著相关性(见图4)。此外,连续4年干旱处理下,植物群落地上净初级生产力与植物群落功能多样性之间没有显著相关性(见图5)。

图 4 极端干旱处理下植物群落性状权重均值与地上净初级生产力的关系
Figure 4. Relationships between aboveground net pramary productivity and community-weighted trait means under extrme drought condition
图 5 极端干旱处理下植物群落性状多样性与地上净初级生产力的关系
Figure 5. Relationships between aboveground net pramary productivity and community trait diversity under extrme drought condition
-
植物功能性状能够反映植物对生长环境的响应和适应,并将环境、植物个体和生态系统结构、过程与功能联系起来的属性或特征,在历史进化过程中,与生存环境相互作用,逐渐形成的形态、生理等方面适应策略[9]。大量研究结果表明,极端干旱可以显著改变草原植物物种水平的性状分布模式。例如,区域尺度上植物比叶面积与土壤水分有效性之间呈显著正相关关系,表明植物养分利用效率与水分利用效率之间存在权衡关系,是植物对资源环境变化的有效适应对策[12]。然而,植物群落水平性状格局变化更加复杂,受种内变异(物种个体性状值变化)和种间变异(物种相对丰度变化)共同调控[5]。以本研究中植物群落高度这一性状为例:极端干旱干扰后,植物群落中大部分物种的个体高度显著降低(种内性状变异);同时,群落结构和组成发生明显变化,相对矮小的稀有物种逐渐消失,较为高大的物种所占比例明显增加,从而提高群落高度值(种间性状变异)。此时,植物群落中的种间与种内性状变异之间存在拮抗作用,致使植物群落权重高度均值没有发生显著变化。由此可见,种内和种间性状的作用关系在维持植物群落性状稳定性及其与生态系统功能关系方面发挥着重要作用[5]。植物群落内部物种的资源生态位具有特异性,从而导致物种对干旱胁迫具有不同响应规律,植物群落水平性状对干旱等环境的响应不能完全基于物种水平的方法进行推演,应当同时考虑种内、种间变异以及两者的共同作用。
本研究中,草甸草原植物群落性状组成对极端干旱的响应具有一定保守性,表明草原物种倾向于采取忍耐策略来适应极端干旱环境。Copeland等(2016)[13]的研究结果同样发现,极端干旱对加利福尼亚草本植物群落高度、比叶面积、叶氮含量等性状没有显著影响。2009年,Smith[8]提出了极端干旱事件对植物群落恢复过程影响的理论框架。在干旱胁迫初始阶段,干旱强度弱且时间短,不能对植物群落结构和功能产生显著影响;随干旱继续发展,干旱开始对植物群落中的个体或种群产生瞬时影响,群落可快速恢复到以前状态;随干旱胁迫持续加重,群落物种组成产生了较大变化,植物群落结构和功能的恢复时间明显增加;当干旱胁迫达到一定程度以后,群落结构发生了质的变化,一旦超过响应阈值,某些植物群落将演替至新的稳定状态,不能恢复到干扰以前的原状态。因此,在功能复杂的植物群落中,极端干旱对植物群落性状组成的影响需要更长时间才能显现出来。
大量研究结果表明,群落性状多样性在一定程度上反映物种在资源利用上的生态位互补,可用于探索植物生态系统功能变化潜在生态学机制,与物种多样性相比,群落性状多样性与生态体统功能关系更为密切 [10, 14]。关于植物性状与生态系统功能关系,Tilman[11]提出了经典的“多样性”假说,该假说认为群落水平上多维性状结构组成越复杂,植被在多维生态空间中的配置越多元化,资源配置与利用越合理充分,则越有利于生态系统功能的提高,反之亦然。然而,本研究结果与此相悖,发现草原植物群落性状多样性不能有效解释植物群落初级生产力对干旱事件的响应规律,表明植物性状多样性不是调控草原植物群落生态适应策略的主要影响因素。由于我国内蒙古草原长期处于水分缺乏的自然条件下,在长期的进化过程中,植物形成了适应干旱生境的逆境属性,致使植物群落性状均值在本研究中成为调控草原初级生产力应对干旱环境的主要驱动因素。该结果与Grime[15]提出的“质量比”假说相一致,认为植物群落优势植物的功能属性是生态系统功能的主要驱动力,强调了优势物种在维持生态系统功能方面的重要作用。
水分和养分是草地植物生长、发育的主要限制因子,所以我们预想,水分胁迫条件下优势植物应当优先优化水分利用性状(如比叶面积、干物质含量)和养分利用性状(如叶氮、叶磷含量),调整植物初级生产力对极端干旱的响应。然而,本研究发现,在极端干旱环境中植物群落光合利用性状(如群落高度、叶碳含量)与植物初级生产力的关系最为密切,该结果与我们的预测不一致。植物高度是直接反映植物群落资源利用策略的综合性指标,干旱环境中植物群落高度越高,地上净初级生产力越高[16]。干旱胁迫下,植物会主动形成更多的单宁等防御性物质,该类物质富含结构性碳,同时,植物生长速率显著降低,导致植物群落净初级生产力与群落叶碳含量呈显著负相关关系[12]。这些研究表明,在草地资源的管理和利用过程中要重视优势物种的功能属性在维持草地群落功能及稳定性过程中的重要作用。本研究结果表明尽管植物群落性状自身对极端干旱的响应不显著,但其在调控草原生态系统功能方面仍发挥着非常重要的作用。Suding[17]提出的功能响应-效应性状理论认为,物种通过改变响应性状来适应环境变化,通过优化效应性状以调控生态系统功能。因此,本研究中观测的植物群落水平性状都不是响应性状,而植物群落高度和叶碳含量是效应性状。未来研究需要分析响应性状和效应性状之间的关联性,探究植物群落响应性状如何影响效应性状以影响生态系统的整体功能[17-18]。
-
近几十年来,极端干旱事件频发对全球和区域尺度上的草地生态系统和人类社会都带来了很大影响。本研究通过控制野外降水,模拟极端干旱事件,探究草甸草原植物群落性状组成以及与生态系统功能的关系对极端干旱的响应,发现植物群落性状对极端干旱的响应具有保守性,植物采用忍耐型对策来适应干旱环境。与多样性假说相悖,群落性状多样性与生态系统功能之间不存在显著关系。相反,与质量比假说相一致,植物群落权重性状均值与生态系统功能之间的关系更为密切,强调了群落中优势物种性状在生态系统功能维持和提升发面的重要地位。本研究结果有助于揭示极端干旱对植物群落结构和功能影响的生态学机制,为草地生态系统植被的保护提供科学依据。
Responses of Plant Community Traits to Extreme Drought in a Meadow Grassland, Inner Mongolia
More Information-
摘要: 人类活动显著影响着全球大气循环格局,导致极端干旱事件发生幅度、频度和持续时间不断增加,这对草原生态系统产生了更多的负面影响。本研究以内蒙古草甸草原植物群落为研究对象,通过连续4年调控生长季降雨的干旱模拟实验,探究植物群落性状组成对干旱胁迫的响应规律及其对生态系统功能的影响。结果表明:极端干旱事件对植物群落性状组成的影响不显著,表明植物群落性状具有一定保守性,植物采用忍耐型对策来适应干旱环境。植物群落地上净初级生产力与群落性状多样性之间关系不显著,该结果不支持多样性假说。植物群落地上净初级生产力与植物群落权重性状均值之间的关系更为密切,与高度显著正相关,与叶碳含量显著负相关,强调了群落中优势物种性状在生态系统功能提升方面的重要地位,该结果支持质量比假说。本研究结果对开展全球变化背景下草原生态系统结构和功能对极端干旱事件的响应机制具有重要指导意义,为草地植被的科学保护和管理提供理论依据。Abstract: Human activities exert a significant impact on the global atmospheric circulation pattern, resulting in the increasing amplitude, frequency and duration of extreme drought events, which have more negative impacts on grassland ecosystem. In this study, we conducted a four-year extreme drought experiment to investigate the responses of plant community traits to extreme drought as well their consequences on ecosystem functions in a meadow grassland, Inner Mongolia. The results showed that experimental extreme drought exerted no significant effect on plant community traits, which indicated that plant are insensitive to extreme drought and plant community adopted tolerance/conservative strategy under water stress conditions. The relationships between plant community trait diversity and aboveground net primary productivity were not observed under extreme drought conditions, which was inconsistent with the diversity hypothesis. On the other hand, aboveground net primary productivity was mediated by the community-weighted trait means under extreme drought conditions. Specifically, aboveground net primary productivity was positively correlated with community-weighted plant height and negatively correlated with community-weighted leaf carbon concentrations. These results highlighted the great effects of dominated species on ecosystem functions, being in line with mass ratio hypothesis. Overall, these results are of great significance for revealing the processes and mechanisms underlying the functional responses of grassland communities to extreme drought, providing a scientific basis for vegetation protection and management in grassland ecosystem.
-
Key words:
- Extreme drought;
- Plant traits;
- Trait diversity;
- Dominant species;
- Adaptive strategy
-
图 2 极端干旱对植物群落性状权重均值的影响
Fig. 2 Effects of extreme drought on plant community-weighted trait means
图 3 极端干旱对植物群落性状多样性的影响
Fig. 3 Effects of extreme drought on plant community trait diverity
图 4 极端干旱处理下植物群落性状权重均值与地上净初级生产力的关系
Fig. 4 Relationships between aboveground net pramary productivity and community-weighted trait means under extrme drought condition
-
[1] Kang, L. , Han, X. , Zhang, Z. & Sun, O. J. . Grassland ecosystems in China: review of current knowledge and research advancement. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 2007, 362: 997−1008. [2] IPCC. Climate change. The physical and science basis. Summary for the policymakers, technical summary and frequently asked questions. Part of the I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC, Geneva, Switzerland. 2021. [3] Jentsch, A. , Kreyling, J. & Beierkuhnlein, C. . A new generation of climate-change experiments: events, not trends. Frontiers in Ecology and the Environment, 2007, 5: 365−374. [4] 张彬, 朱建军, 刘华民, 等. 极端降水和极端干旱事件对草原生态系统的影响. 植物生态学报 2014, 38: 1008−1018. [5] Jung, V. , Albert, C. H. , Violle, C. , Kunstler, G. , Loucougaray, G. & Spiegelberger, T. . Intraspecific trait variability mediates the response of subalpine grassland communities to extreme drought events. Journal of Ecology, 2014, 102: 45−53. [6] 宋琳, 雒文涛, 马望, 等. 极端干旱事件对草甸优势草原植物非结构性碳水化合物的影响, 植物生态学报, 2020, 44: 669−676. [7] Chen LP, Zhao NX, Zhang LH, Gao YB. Responses of two dominant plant species to drought stress and defoliation in the Inner Mongolia Steppe of China. Plant Ecology, 2013, 214: 221–229. [8] Smith MD et al. . A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change. Ecology . 2009, 90: 3279−3289. [9] He, N. , Liu, C. , Piao, S. , Sack, L. , Xu, L. , Luo, Y. et al. . Ecosystem Traits Linking Functional Traits to Macroecology. Trends in Ecology & Evolution, 2019, 34: 200−210. [10] McGill, B. J. , Enquist, B. J. , Weiher, E. & Westoby, M. . Rebuilding community ecology from functional traits. Trends in Ecology & Evolution, 2006, 21: 178−185. [11] Tilman D. Plant Strategies and the Dynamics and Structure of Plant Communities. . Princeton University Press, Princeton, NJ. 1988. [12] Wright IJ et al. The worldwide leaf economics spectrum. Nature, 2004, 428: 821−827. [13] Copeland, S. M. , Harrison, S. P. , Latimer, A. M. , Damschen, E. I. , Eskelinen, A. M. , Fernandez-Going, B. et al. . Ecological effects of extreme drought on Californian herbaceous plant communities. Ecological Monographs, 2016, 86: 295−311. [14] Luo, W. , Griffin-Nolan, R. J. , Ma, W. , Liu, B. , Zuo, X. , Xu, C. et al. . Plant traits and soil fertility mediate productivity losses under extreme drought in C3 grasslands. Ecology, e03465. 2021. [15] Grime JP. Benefits of plant diversity to ecosystems: immediate, filter and founder effects. Journal of Ecology, 1998, 86: 902−910 [16] Falster, D. S. , Westoby, M. . Plant height and evolutionary games. Trends in Ecology & Evolution, 2003, 18: 337−343. [17] Suding, K. N. , Lavorel, S. , Chapin, F. S. , III, Cornelissen, J. H. C. , Diaz, S. , Garnier, E. et al. . Scaling environmental change through the community-level: a trait-based response-and-effect framework for plants. Glob. Change Biol. , 2008, 14: 1125−1140. [18] 李瑞新, 丁勇, 马文静, 等. 植物功能多样性及其与生态系统功能关系研究进展. 生态环境学报, 2016, 25: 1069−1075. -

 点击查看大图
点击查看大图

图(5)
计量
- 文章访问数: 544
- HTML全文浏览量: 267
- PDF下载量: 111
- 被引次数: 0
















