










 下载:
下载:

-
动物行为是动物个体或社群适应内外环境所作的反应,是对周围的生物和非生物环境动态的一种适应[1]。在行为生态学研究中,日活动节律指动物在一天中不同时间段的活动强度及其周期性变化规律,主要由动物生物钟控制[2],同时还受到其他因素的调节,如环境温度及其季节的变化[3, 4]、食物可获得性[5]、内源节律[6]、种间关系[7]等,因此动物会改变其活动模式,以平衡收益与成本[8]。
在温带地区,光照周期和昼夜长短会随着季节变化而变化,从而影响食草动物的生理和行为[9]。比如,高温会导致白面牛羚 (Damaliscus dorcas phillipsi) 白天活动减少,夜间活动相应增加[10];低温会迫使羱羊 (Capra ibex) 冬季新陈代谢下降,以较少热量消耗以便顺利越冬[11]。所以,动物的活动节律会随季节进行相应的调整[5],活动节律是动物本身适应进化和应对环境因素变化而形成的生存策略[12]。
毛冠鹿 (Elaphodus cephalophus) 在我国主要分布于南方山地丘陵地带,食性杂,对环境变化较为敏感[13]。毛冠鹿生性胆小,倾向于选择较为隐蔽的生境[14],因此选择具干扰小、持续时间长、全天候检测、数据和观察更加客观和稳定、受天气影响小等优点的红外相机技术,在研究动物活动节律方面得到了广泛应用[12,15-20]。
山地环境具有湿度高、温度高、地形复杂等特点[21],加上植被的垂直地带性和季节变化,意味着资源与环境的季节间波动。因此,我们认为毛冠鹿在不同季节会为了适应环境变化而有不同的行为策略,日活动模式也呈现出明显的季节变化。然而,至今尚未见关于毛冠鹿行为节律和季节变化方面的专题报道。基于此,本研究选择四川白河国家级自然保护区 (以下简称“白河”) 毛冠鹿种群作为研究对象,利用红外相机技术获取调查区域内毛冠鹿的影像数据,分析其活动节律分布、影响因素和季节变化,以期为毛冠鹿种群的管理和保护提供基础数据支持。
-
白河位于四川省阿坝藏族羌族自治州九寨沟县境内 (104°0'—104°12' E,33°10'—33°22' N),属岷山山系北段,是青藏高原向四川盆地的过渡地带,最高海拔4453 m,最低海拔1 240 m,总面积162 km2,其中林地157.57 km2,占总面积的89%[22]。保护区属暖温带半湿润气候,多年平均气温11.5 ℃,7月平均气温20 ℃,1月平均气温–0.5 ℃,多年平均降水量约610 mm[23]。区内植被类型具有明显的垂直分异,随海拔变化依次为落叶阔叶林、针阔混交林、亚高山针叶林和高山草甸[22]。四川羚牛(Budorcas tibetanus)、林麝(Moschus berezovskii)、马麝(Moschus sifanicus)、中华斑羚(Naemorhedus griseus)、中华鬣羚(Capricornis milneedwardsii)、毛冠鹿等有蹄类在白河均有分布[24],但尚未发现雪豹(Panthera uncia)等大中型食肉动物的分布。
-
在保护区及周边地区共布设红外相机调查位点160个(保护区内122个,保护区周边地区38个),所有红外相机监测均按照沿样线海拔每上升100 m安放1台红外相机的原则布设,覆盖海拔范围为1700~3700 m,红外相机的具体设置标准见[25]。
-
基于独立探测照片计算毛冠鹿的活动强度,独立有效照片 (independent photo,IP) 的定义为30 min内连续多次拍到的同一物种的所有图像记为1张IP。季节按照春 (3—5月)、夏 (6—8月)、秋 (9—11月)、冬 (12—翌年2月) 设置,并将6:00—20:00时定为昼间,18:00—6:00时定为夜间。本研究以1小时为间隔划分时间段,即00:00—01:00时记为1,01:00—02:00时记为2,以此类推,共计24个时间段,并以此作为统计IP数的依据,用于计算各个时间段的相对活动强度指数 (relative activity index)[26],相对活动强度指数=本时间段的有效探测数/总的有效探测数。
利用核密度函数建立毛冠鹿日活动节律的函数模型[27],该方法认为物种的每次探测是从连续的日活动节律分布中采集的随机样本,这个日活动节律分布描了该物种在某个特定时间段被探测到的概率。核密度函数公式为:
$$ \widehat{f}\left(x\right)=\frac{1}{n}{\sum }_{i=1}^{n}{k}_{v}\left[d\left(x,{x}_{i}\right)\right] $$ (1) 式中,KV为vonMises分布的概率密度函数,d(x,xi)为任意一点x与样本量xi之间的角度距离。同时,采用重叠指数 (coefficient of overlap)[27]计算毛冠鹿的日活动节律在不同季节间的重叠程度,用不同季节日活动节律分布曲线重叠的面积比
$ \widehat{\Delta } $ 表示:0表示完全分离,1表示重叠。所有分析均在R4.0.0软件中进行,使用的分析包为overlap。以18:00—06:00作为夜行性分析时间段,以2 h为时间间隔,计算夜间相对丰富度(night- time relative abundance index,NRAI),依据毛冠鹿活动时间段分布比例,结合公式分析其夜行性[19]:
$$ {{\rm{I}}_{NRAI}} = D/N*100{\text \%} $$ (2) 式中,D为毛冠鹿在夜间时间段的独立有效照片数,N为毛冠鹿的独立有效照片总数。若NRAI值>13/24,则其具有夜行性,反之则具有昼行性,接近则认为夜行性不明显。
-
在野外布设相机陷阱调查位点160个,除2个位点由于相机故障而未能获得有效数据外,共有123台相机拍摄到了毛冠鹿,剔除掉IP少于10的相机位点,共有55个相机位点,共拍摄到毛冠鹿独立照片1 069张。
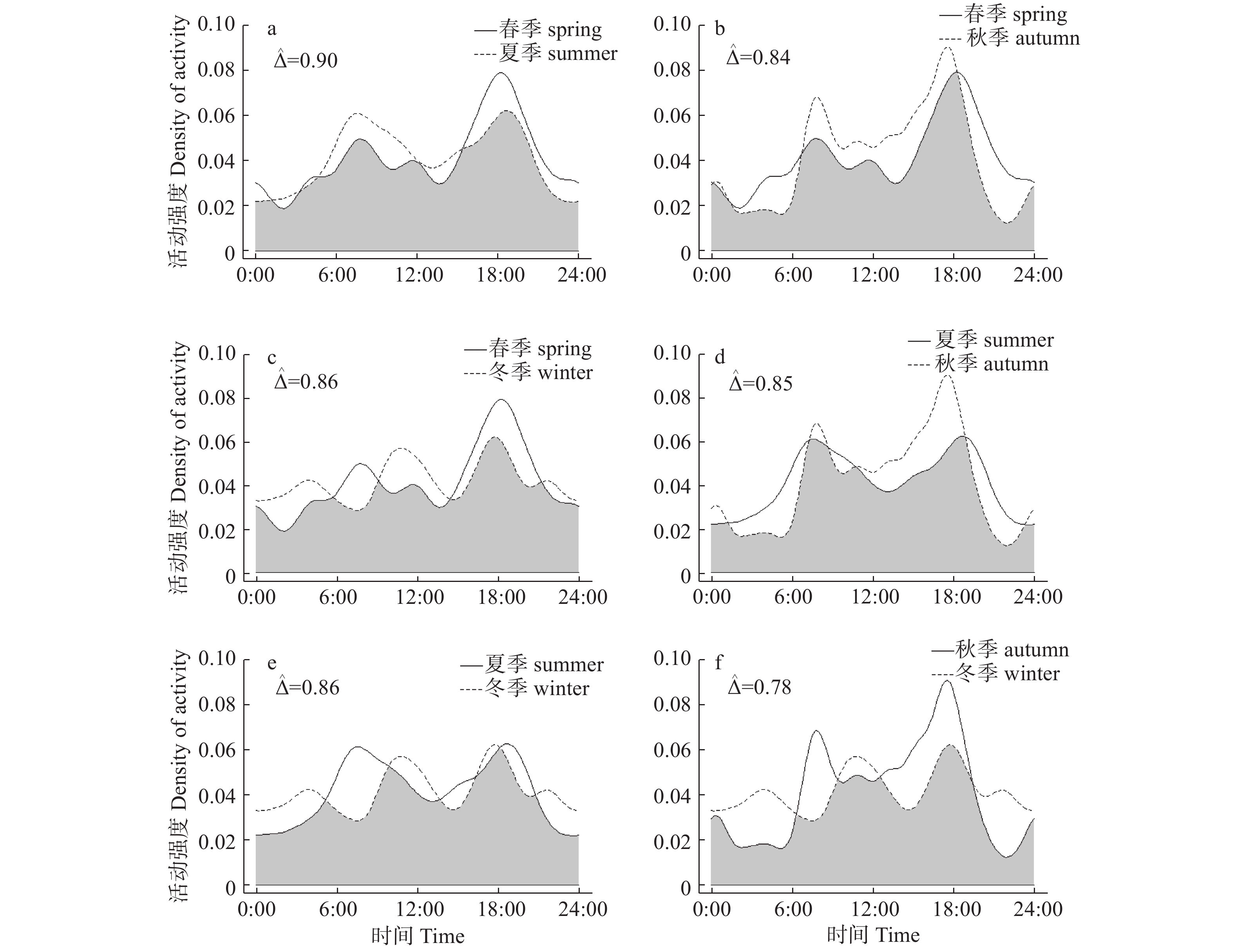
根据红外相机数据,毛冠鹿全年的日活动呈现明显的双峰趋势(见图1),主要在晨昏前后活动。6:00—10:00时,16:00—20:00时毛冠鹿活动强度最高,1:00—3:00时活动强度最低。
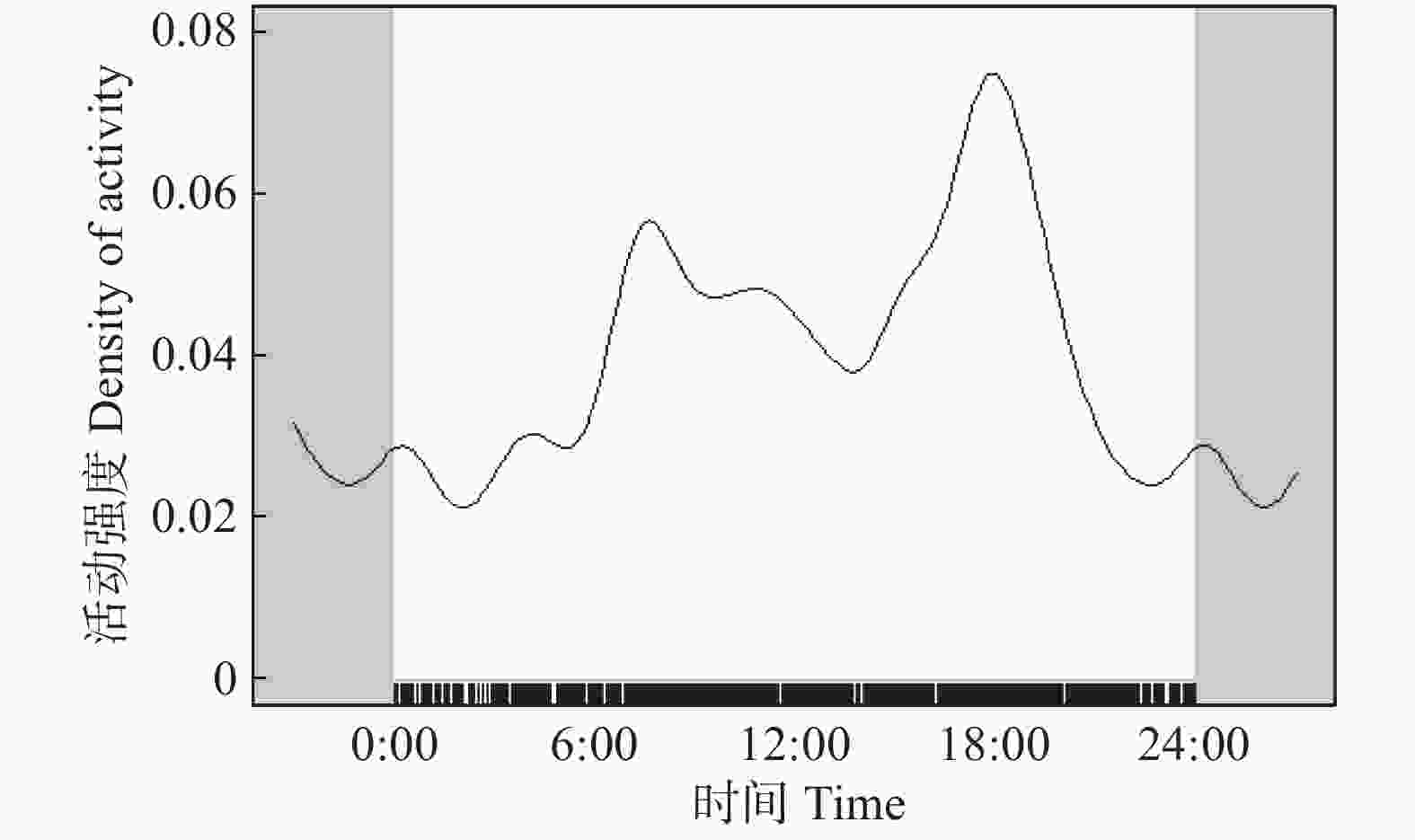
图 1 毛冠鹿全年日活动节律
Figure 1. Annual daily activity rhythm of Elaphodus cephalophus
-
在四个季节(春季、夏季、秋季、冬季)拍摄到毛冠鹿独立照片数分别为202、336、320、211。毛冠鹿日活动节律的重叠指数在不同季节间均较高,其中春季和夏季重叠指数最高(
$ \widehat{\Delta }=0.90 $ ),秋季和冬季重叠指数最低($ \widehat{\Delta }=0.78 $ )(见图2)。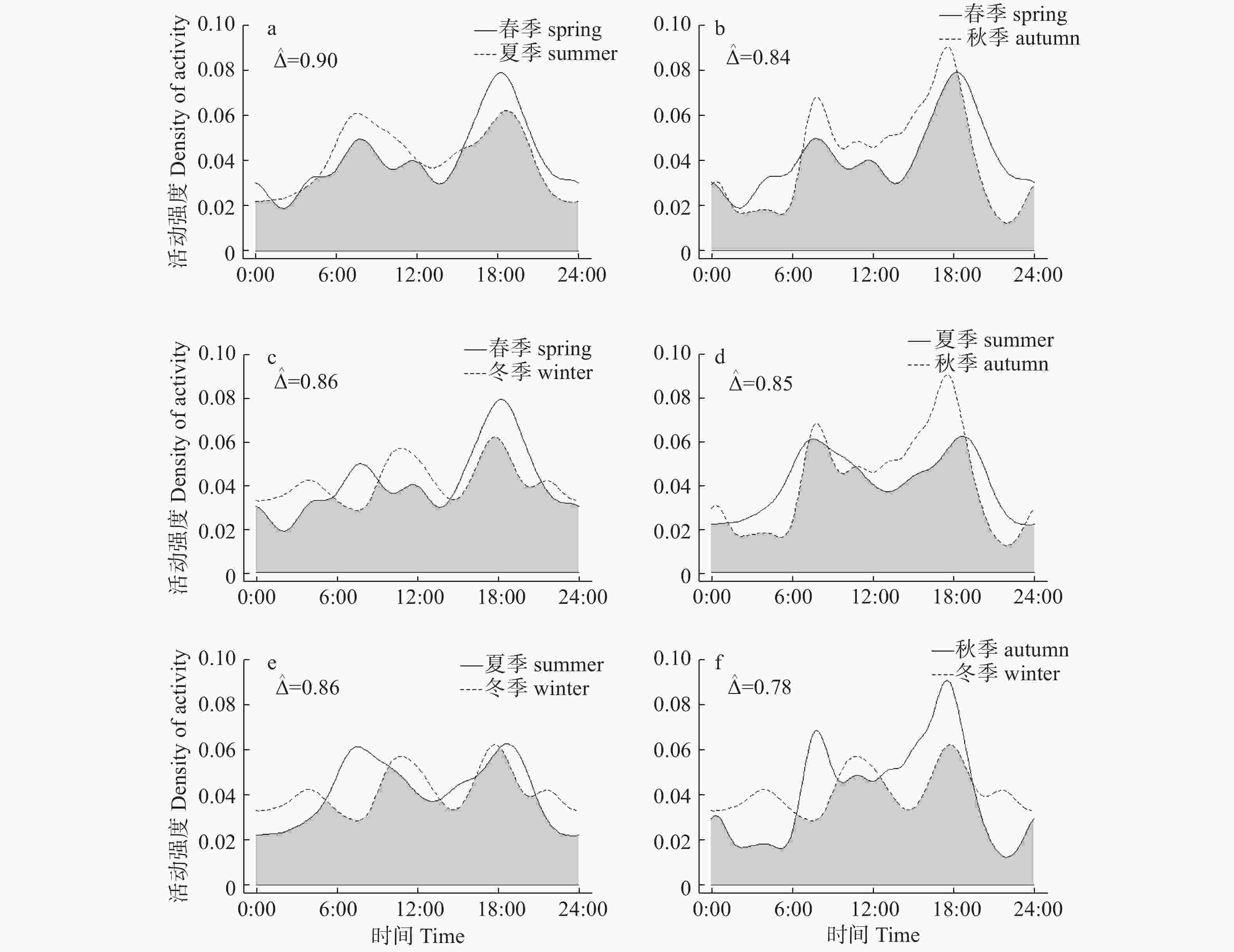
图 2 毛冠鹿不同季节的日活动节律比较
Figure 2. Comparison of daily activity rhythms of Elaphodus cephalophus in different seasons
季节间的比较的结果表明,毛冠鹿日活动节律存在季节性变化,春季和秋季活动模式相似(见图2b),一天中有两个活动较高的时段(春季:6:00—8:00时、18:00—20:00时;秋季:6:00—8:00时、16:00—18:00时),在10:00—14:00时活动强度相对较低;进入秋季以后,日活动时间开始缩短(见图2b),在冬季达到最低:早上活动高峰推迟至8:00时开始,傍晚活动高峰结束时间相较于夏季提前至18:00时(见图2e)。
-
在18:00—6:00时间段独立照片数分别为427,夜间相对丰富度为39.94%,低于13/24,不具有明显的夜行性,表明毛冠鹿的活动主要集中在白昼。春夏秋冬夜间独立照片数分别96、134、96、101,夜间相对丰富度分别为47.52%、39.88%、30%、47.87%。
-
昼夜活动节律是动物对环境条件的高度适应,受到季节、温度、性别、年龄、繁殖周期和光周期等诸多因素的影响[10],它与动物体的代谢和能量有关,而且会随着生态条件的变化而发生改变[28],不同的物种其主导因素不同[29],一般认为,昼夜更替为活动节律的决定因素,食物、能量、性别、遗传、繁殖状况、种间竞争、社群以及天气状况等为主导因素[30]。
研究表明,许多有蹄类的日活动行为格局相对比较固定,具有晨昏活动的节律[31],如水鹿 (Rusa unicolor)[20]、小麂(Muntiacus reevesi)[32]、黑麂 (Muntiacus crinifron)[15]、林麝 (Moschus berezovskii)[33]、非洲水牛(Syncerus caffer)[34]等的日活动高峰都出现在当地季节性日出日落前后。本研究发现,与其他温带有蹄类一样,毛冠鹿存在相似的晨昏活动节律,在全年呈现明显的双峰活动模式(见图1),昼间呈现出取食-休息-取食规律。在不同季节,日活动节律与全年一致,存在一定的季节变化。与贾晓东等[33]和孙佳欣等[19]对于毛冠鹿的研究一致。与于桂清[35]和穆君等[36]研究不同,造成此差异的原因有待进一步分析。
毛冠鹿的日活动节律呈现明显的季节性差异,我们分析认为气候条件和食物资源是其日活动节律模式及季节性变化的主要驱动因素。毛冠鹿选择在晨昏活动的活动模式,一方面有利于避开不利气候条件的胁迫,因为在午间光照强度大,另一方面毛冠鹿作为反刍动物,在早上经过几个小时的取食,会进行一段时间的消化。春冬季相对于夏秋季来说,早上的活动高峰会推迟,傍晚的活动高峰则会提前,同时春冬季的夜间活动强度明显强于夏秋季,我们认为这种变化与恶劣的气候、食物的匮乏有关。冬春季气候寒冷,积雪厚度大,不利于其取食和活动,加之食物质量差,又需增加觅食时间来达到能量需求。因此,我们认为白河毛冠鹿在长期的生存适应过程中,平衡环境条件的变化,在不同季节产生相适应的日活动节律策略,这也表明物种的活动节律有着较强的环境可塑性。
毛冠鹿是广泛分布在西南山地森林生态系统中的优势有蹄类[19]。鉴于山地环境对气候变化的敏感性,山地有蹄类在生态系统的作用将面临严峻的考验。本研究首次专题报道毛冠鹿的日活动特征及全年各季节变化,为后续研究提供的重要的基础资料,也为保护区红外相机数据的应用方向提供了范例。
Activity Rhythm and Seasonal Changes of Elaphodus cephalophus in Baihe National Nature Reserve, Sichuan Province
More Information-
摘要: 活动节律是动物内源节律应对环境压力的动态生活策略,反映了个体对资源与威胁的适应。毛冠鹿是广泛分布在西南山地森林中的优势有蹄类,但至今未见专题报道。2017年1月至2019年11月,笔者利用红外相机在四川白河国家级自然保护区对毛冠鹿 (Elaphodus cephalophus) 进行了调查,分析了它们的活动节律及其季节性变化。结果显示:1) 毛冠鹿具有明显的晨昏活动,分别在6:00—10:00时,16:00—20:00时活动强度最高;2) 毛冠鹿的活动节律存在季节性变化,春季和夏季重叠指数最高 (
$ \widehat{\Delta }=0.90 $ ),秋季和冬季重叠指数最低 ($ \widehat{\Delta }=0.78 $ );春冬季相对于夏秋季,早上的活动高峰推迟,傍晚的活动高峰提前;3) 毛冠鹿不具有明显的夜行性,但冬春季夜间活动明显高于夏秋季。我们认为极端温度和食物供给是其日活动节律季节变化的主要驱动因素。#共同第一作者Abstract: Activity rhythm is a dynamic life strategy of animal endogenous rhythm in response to environmental stress, which reflects the individual's adaptation to resources and threats. Tufted deer (Elaphodus cephalophus) is the dominant ungulate species widely distributed in southwest mountain forests of China, but no special reports have been published on it so far. From January 2017 to November 2019, infrared cameras were used to investigate the Tufted deers in Baihe National Nature Reserve in Sichuan province, in order to reveal their activity rhythm and seasonal changes. The results showed that: 1) Tufted deer had obvious typical dawn and dusk activity, with the highest activity intensity from 6:00 to 10:00 and from 16:00 to 20:00 respectively. 2) The seasonal changes of activity rhythm was apparent, with the highest overlapping index in spring and summer ($ \widehat{\Delta }=0.90 $ ) and the lowest overlapping index in autumn and winter ($ \widehat{\Delta }=0.78 $ ). Compared with summer and autumn, the activity peak in the morning was delayed and the activity peak in the evening was advanced in spring and winter. 3) Tufted deer had no obvious nocturnal activity, but the nocturnal activity in winter and spring was obviously higher than that in summer and autumn. We believed that extreme temperature and food supply were the main drivers of seasonal variations of their daily activity rhythms.-
Key words:
- Ungulate;
- Mountain forest;
- Southwest mountains;
- Activity rhythm
-
[1] 孙儒泳. 动物生态学原理 第3版[M]. 北京: 北京师范大学出版社, 2001. [2] 尚玉昌. 动物的行为节律[J]. 生物学通报,2006,41(10):8−10. doi: 10.3969/j.issn.0006-3193.2006.10.004 [3] P Beier, Mccullough D-R. Factors influencing white-tailed deer activity patterns and habitat use[J]. Wildlife Monographs, 1990, 109(109): 3−51. [4] Garrett Street, Rodgers Art, Fryxell John. Mid-Day Temperature Variation Influences Seasonal Habitat Selection by Moose[J]. Journal of Wildlife Management, 2015, 79(3): 505−512. doi: 10.1002/jwmg.859 [5] Ariane Massé, Cté Steeve-D. Spatiotemporal variations in resources affect activity and movement patterns of white-tailed deer (Odocoileus virginianus) at high density[J]. Canadian Journal of Zoology, 2013, 91(4): 252−263. doi: 10.1139/cjz-2012-0297 [6] Gabriel Hofmann, Coelho Igor, Bastazini Vinicius, et al. Implications of climatic seasonality on activity patterns and resource use by sympatric peccaries in northern Pantanal[J]. International Journal of Biometeorology, 2016, 60(3): 421−433. doi: 10.1007/s00484-015-1040-8 [7] Mathias Tobler, Carrillo-Percastegui Samia, Powell George. Habitat use, activity patterns and use of mineral licks by five species of ungulate in South-Eastern Peru[J]. Journal of Tropical Ecology, 2009, 25(3): 261−270. doi: 10.1017/S0266467409005896 [8] Marcus Rowcliffe, Kays Roland, Kranstauber Bart, et al. Quantifying levels of animal activity using camera-trap data[J]. Methods in Ecology and Evolution, 2014, 5(11): 1170−1179. doi: 10.1111/2041-210X.12278 [9] Barry Lovegrove. Seasonal thermoregulatory responses in mammals[J]. Journal of comparative physiology. B, Biochemical, systemic, and environmental physiology, 2005, 175(4): 231−247. doi: 10.1007/s00360-005-0477-1 [10] Klaus-M Scheibe, Robinson Tracy-L, Scheibe Annemarie, et al. Variation of the phase of the 24-h activity period in different large herbivore species under European and African conditions[J]. Biological Rhythm Research, 2009, 40(2): 169−179. doi: 10.1080/09291010701875070 [11] Claudio Signer, Ruf Thomas, Arnold Walter. Hypometabolism and basking: the strategies of Alpine ibex to endure harsh over-wintering conditions[J]. Functional Ecology, 2011, 25(3): 537−547. doi: 10.1111/j.1365-2435.2010.01806.x [12] 刘鹏,刘振生,高惠,等. 基于红外相机技术的贺兰山同域分布阿拉善马鹿和岩羊活动规律研究[J]. 生态学报,2019,39(24):9365−9372. [13] 贾晓东. 秦岭南坡不同干扰机制下的有蹄类动物多样性研究[D]. 西北大学, 2015. [14] 岳建兵. 4种有蹄类动物栖息地选择比较研究[J]. 林业资源管理,2014,6(3):109−112. [15] 章书声,鲍毅新,王艳妮,等. 基于红外相机技术的黑麂活动节律[J]. 兽类学报,2012,32(4):368−372. [16] 李晟,王大军,肖治术,等. 红外相机技术在我国野生动物研究与保护中的应用与前景[J]. 生物多样性,2014,22(6):685−695. [17] 黄沛琳,肖文宏,杨海涛,等. 东北梅花鹿种群活动节律和集群行为研究[J]. 北京师范大学学报(自然科学版),2015,51(5):498−503. [18] 吴兵,初雯雯,吴洪潘,等. 卡拉麦里山有蹄类自然保护区水源地野放普氏野马的活动节律: 基于红外相机监测数据[J]. 动物学杂志,2017,52(4):545−554. [19] 孙佳欣,李佳琦,万雅琼,等. 四川9种有蹄类动物夏秋季活动节律研究[J]. 生态与农村环境学报,2018,34(11):1003−1009. doi: 10.11934/j.issn.1673-4831.2018.11.007 [20] 何兴成,付强,吴永杰,等. 水鹿的群体结构和活动节律分析[J]. 兽类学报,2019,39(2):134−141. [21] 张荣祖. 中国动物地理[M]. 北京: 科学出版社, 2011. [22] 金贵祥,李沂韦,孙治宇,等. 白河国家级自然保护区川金丝猴的分布和数量[J]. 普洱学院学报,2020,36(3):17−21. doi: 10.3969/j.issn.2095-7734.2020.03.006 [23] 李艳忠,董鑫,刘雪华. 40年岷山地区白河自然保护区川金丝猴的生境格局动态[J]. 生态学报,2016,36(7):1803−1814. [24] 郭家强. 岷山山系两个川金丝猴种群间活动节律与活动时间分配的比较[D]. 西华师范大学, 2018. [25] 陈星,赵联军,胡茜茜,等. 基于地形的牲畜空间利用特征及干扰评价——以王朗国家级自然保护区为例[J]. 生物多样性,2019,27(6):630−637. [26] Sheng Li, McShea William, Wang Dajun, et al. The use of infrared‐triggered cameras for surveying phasianids in Sichuan Province, China[J]. Ibis, 2010: 152299−309. [27] Martin Ridout, Linkie Matthew. Estimating Overlap of Daily Activity Patterns From Camera Trap Data[J]. Journal of Agricultural Biological and Environmental Statistics, 2009: 14322−337. [28] 刘国库,周材权,杨志松,等. 竹巴笼矮岩羊昼间行为节律和时间分配[J]. 生态学报,2011(04):972−981. [29] Arantza Aldezabal, Garin I, García-González Ricardo. Activity rhythms and the influence of some environmental variables on summer ungulate behaviour in Ordesa-Monte Perdido National Park[J]. Pirineos: Revista de Ecología de Montaña, 1999(153): 145−157. [30] 宋玉成,赵蕙,时磊. 吐鲁番沙虎的活动节律及其影响因子分析[J]. 新疆农业大学学报,2009,32(1):22−25. doi: 10.3969/j.issn.1007-8614.2009.01.005 [31] 连新明,李晓晓,颜培实,等. 夏季可可西里雌性藏原羚行为时间分配及活动节律[J]. 生态学报,2012(03):663−670. [32] 官天培,龚明昊,胡婧,等. 小麂秋季利用人工盐场的节律[J]. 动物学杂志,2015,50(2):169−175. [33] 贾晓东,刘雪华,杨兴中,等. 利用红外相机技术分析秦岭有蹄类动物活动节律的季节性差异[J]. 生物多样性,2014,22(6):737−745. [34] N Owen Smith, Goodall V. Coping with savanna seasonality: Comparative daily activity patterns of African ungulates as revealed by GPS telemetry[J]. Journal of Zoology, 2014, 293(3): 181−191. doi: 10.1111/jzo.12132 [35] 于桂清,康祖杰,刘美斯,等. 利用红外相机对湖南壶瓶山国家级自然保护区兽类和鸟类多样性的初步调查[J]. 兽类学报,2018,38(1):104−112. [36] 穆君,王娇娇,张雷,等. 贵州习水国家级自然保护区红外相机鸟兽监测及活动节律分析[J]. 生物多样性,2019,27(6):683−688. -

 点击查看大图
点击查看大图
图(2)
计量
- 文章访问数: 596
- HTML全文浏览量: 257
- PDF下载量: 42
- 被引次数: 0














