






 下载:
下载:

-
雄黄桂(Osmanthus fragrans 'Xionghuang')是木犀科木犀属的常绿小乔木,属于秋季桂丹桂品种群,这种植物以其高度的观赏价值而备受青睐,此外,雄黄桂发达的根系和抗污染能力使其在生态修复和土壤保护方面发挥着积极作用,因此在城市绿化[1]、生态保护等方面具有一定的意义和价值。雄黄桂原产于中国南方地区,虽然目前已在多地进行了栽培,但相对于一般的桂花来说,雄黄桂的生长速度较慢,这可能导致景观效果延迟、观赏价值下降、生态服务功能降低等,因此这一问题亟待解决。
林下植被在人工林中扮演着重要的角色,是人工林的重要组成部分[2]。过去的多数观点认为,林下植被与上层乔木存在着竞争,涉及到水分[3]、养分[4]和生存空间[5]等方面的问题。然而,现有的研究显示,林下植被也具有部分积极的生态功能,它有助于提高土壤含水率[6-7]、降低土壤容重[8],同时改善土壤微环境,这些效应共同改变了林地土壤的养分循环和供应。除此之外,林下植被还可以起到调节乔木层的温度和湿度、抑制杂草的作用,从而减少杂草对乔木层的竞争。光合作用是植物将光能转化为化学能的过程,是植物生长和发育的基础[9],因而研究植物光合作用对不同灌木种类的变化特征对于探讨最适雄黄桂生长的灌木种类具有重要的意义。目前,关于雄黄桂的研究较少,主要集中在花期早晚[10]等方面;关于对生长和光合作用的影响的研究主要集中于施肥[11]、土壤类型[12-13]以及光强[14-15]等,而关于灌木种类的研究尚未见报道。因此,选取了栽植4种不同的灌木的雄黄桂林和空白对照的雄黄桂林设置样地,对雄黄桂的枝条生长和光合作用进行研究,目的是(1)揭示下层灌木对雄黄桂生长和光合作用的影响;(2)揭示雄黄桂枝生长量与光合指标的相关性;(3)选出促进雄黄桂生长最优的灌木,从而为解决雄黄桂生长速度相对较慢的问题提供理论依据。
-
研究区位于四川省成都市新都区(104° 13′ 12.278′′-104° 13′ 16.489′′ E,30° 47′ 35.588′′-30° 47′ 36.826′′ N)内,该研究区属暖湿亚热带湿润气候区,年平均日照时数为1042-1412 h,地形平坦,平均海拔472 m,年平均气温16.1℃,年极端最高气温为37.1℃,最低为−5.4 ℃,年平均无霜期279 d,年平均降雨量为896.8 mm,降水集中在6-9月份,土壤类型以水稻土为主。研究区内优势乔木为雄黄桂(Osmanthus fragrans ‘Xionghuang’),灌木有红花檵木(Loropetalum chinense var. rubrum),山茶(Ca-mellia japonica ),红枫(Acer palmatum ‘Atropurpureum’),金边黄杨(Euonymus japonicus ‘Aurea-marginatus’)等,雄黄桂与其下灌木于2013年同时栽植,栽植后林木自然生长,无人为干扰。
-
2022年10月,按照方精云[16]的研究方法,在对研究区全面踏查的基础上,采用典型样地法选取4种不同的林下灌木和空白对照的雄黄桂林,各设置4个20 ×30 m的标准样地,共15个样地(表1)。每个样地的坡向、坡位及海拔等条件基本一致。在每块样地内沿对角线设置6个5 m×5 m的灌木样方,记录样方内灌木的种名、株数、高度、盖度等。
表 1 样地概况
Table 1. General situation of the sample plot
样地号 灌木种类 平均冠幅(m) 灌木平均树高(m) 平均树高(m) 平均胸径(cm) 郁闭度(%) 密度(plants·hm−2) 1-3 Ⅰ 0.523 0.960 2.34-2.66 7.73-8.59 8-13 825-875 4-6 Ⅱ 1.378 1.322 2.83-2.87 9.12-11.10 17-24 875-975 7-9 Ⅲ 0.959 1.121 2.44-2.67 7.46-8.16 12-19 850-900 10-12 Ⅳ 0.800 1.221 2.51-3.01 8.66-10.93 14-25 900 13-15 CK - - 2.38-2.75 8.45-8.69 12-13 725-875 注:Ⅰ代表茶树,Ⅱ代表红枫,Ⅲ代表红花檵木,Ⅳ代表金边黄杨,CK代表空白对照 -
每株树选取10个长势相似的二级分枝进行编号,以枝的基部为参考点,沿着枝的中心轴线测量到末端。用卷尺(1 cm精度)每隔一周定期测量其长度,最终共测定了8次。
-
在晴天上午9:00-11:30时段,使用Li-6800便携式光合测定仪测定雄黄桂的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)。测定时选取已编号的二级分枝上完全展开的成熟功能叶,每个处理重复测定3个叶片。测定时光强设定为1000 μmol·m−2s−1,CO2浓度设定为400 μmol·mol,叶室温度为30℃。
-
相同测定条件下于9:00—11:30时段,对已编号的二级分枝上生长的叶片进行植物光响应特征参数的测定, 光合光响应特征参数的测定用Li-6800设定叶室内光合有效辐射强度分别为:0、50、100、200、400、600、800、1000、1200、1600、2000、2400、2800 μmol m−2 s−1),并将采用直角双曲线修正模型[16]来拟合光响应曲线。
-
使用Excel、Origin2022软件完成数据整理、制表、绘图,采用SPSS 26.0软件对试验数据进行单因素(one-way ANOVA)方差分析、Pearson相关性分析和 LSD多重比较。
-
根据8次测量的二级枝条长度可计算出每一次测量的枝生长量。如表2和图1可知,Ⅱ和Ⅲ的第一次、第二次和第三次枝生长量极显著大于Ⅰ、Ⅳ和CK,而这三者的第二次和第三次无显著差异;Ⅱ的第四次测量得出的枝生长量极显著大于Ⅰ、Ⅲ、Ⅳ和CK,Ⅲ极显著大于CK,Ⅰ、Ⅳ和CK之间无显著差异;Ⅰ、Ⅱ、Ⅲ、Ⅳ和CK的第五次、第六次和第七次枝生长量均无明显差异。总的来看,Ⅱ的枝生长效果最好,Ⅲ次之,Ⅰ、Ⅳ和CK的效果相当。
表 2 林下栽植不同灌木对雄黄桂枝生长的影响
Table 2. Effects of different shrubs planted under the canopy of Osmanthus fragrans 'Xionghuang' on branch growth of Osmanthus fragrans 'Xionghuang'
灌木种类 枝生长量 1 2 3 4 5 6 7 Ⅰ 5.67±0.25c 1.14±0.06b 1.11±0.27b 0.73±0.15bc 0.38±0.06a 0.18±0.02a 0.12±0.03a Ⅱ 8.59±0.48a 3.68±0.98a 2.27±0.20a 1.37±0.19a 0.45±0.04a 0.26±0.08a 0.15±0.05a Ⅲ 8.47±0.18a 4.47±0.25a 2.11±0.11a 0.89±0.07b 0.42±0.06a 0.28±0.059a 0.13±0.04a Ⅳ 6.63±0.32b 1.47±0.17b 1.26±0.09b 0.72±0.08bc 0.41±010a 0.19±0.03a 0.12±0.02a CK 5.48±0.16c 1.04±0.20b 0.96±0.16b 0.64±0.09c 0.33±0.07a 0.18±0.0a 0.12±0.02a 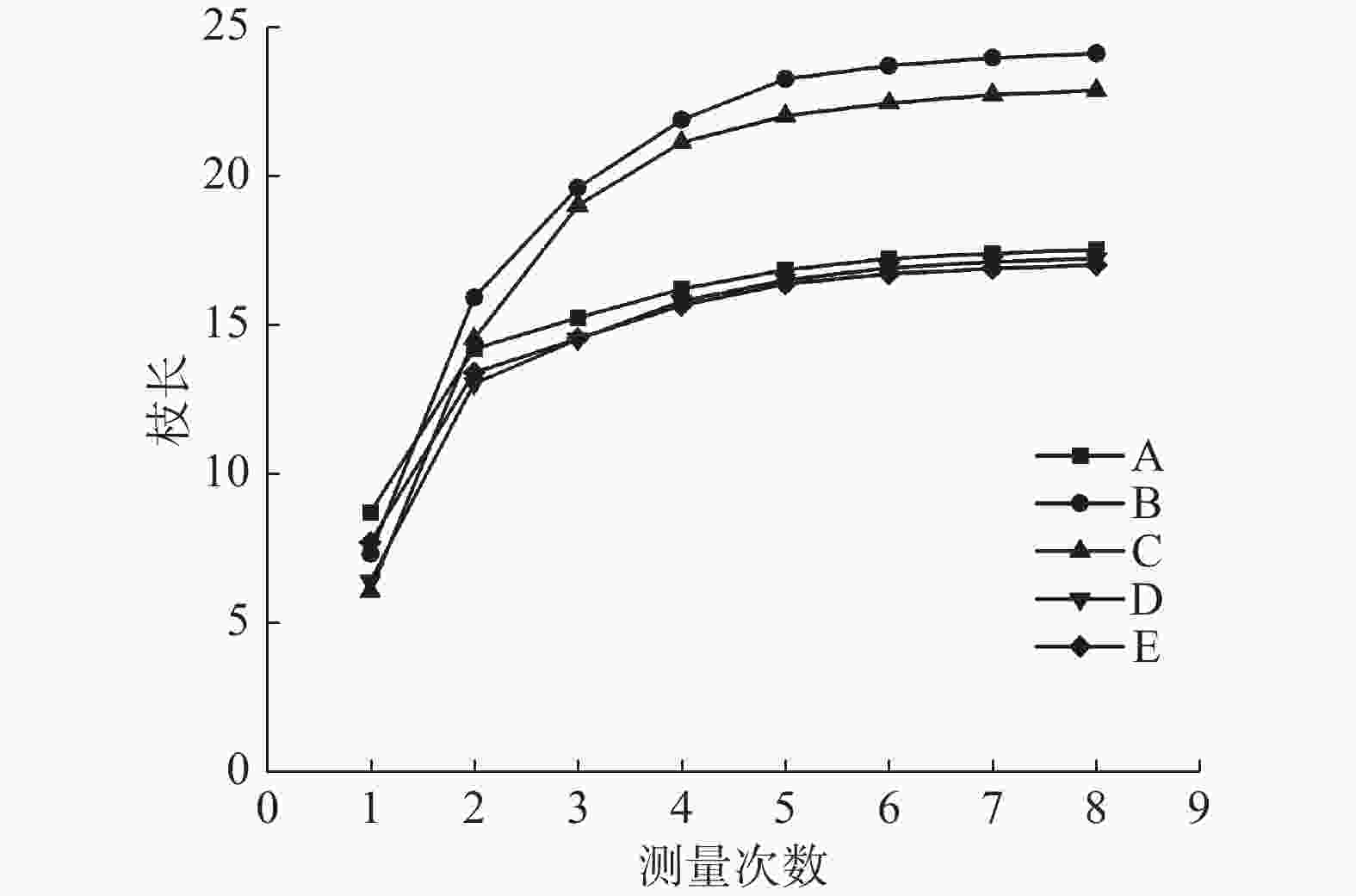
图 1 林下栽植不同灌木对雄黄桂枝生长的影响
Figure 1. Effects of different shrubs planted under the canopy of Osmanthus fragrans 'Xionghuang' on branch growth of Osmanthus fragrans 'Xionghuang'
-
由表3可知,Pn和Gs表现为Ⅱ>Ⅲ>Ⅳ>Ⅰ>CK,Ⅱ的Pn和Gs均与Ⅰ、Ⅳ和CK差异达到极显著水平(P<0.01),而与Ⅲ无显著差异;Ⅲ的Pn和Gs均也与Ⅰ、Ⅳ和CK差异达到极显著水平(P<0.01);Tr也表现为Ⅱ>Ⅲ>Ⅳ>Ⅰ>CK,Ⅱ显著大于Ⅰ、Ⅲ、Ⅳ和CK,Ⅰ、Ⅲ、Ⅳ无显著差异,Ⅲ、Ⅳ显著大于CK,CK和Ⅰ无显著差异;Ci表现为CK>Ⅳ>Ⅰ>Ⅲ>Ⅱ,且Ⅱ、Ⅲ与CK存在极显著差异(P<0.01)。
表 3 林下栽植不同灌木对雄黄桂光合特性的影响
Table 3. Effects of different shrubs planted under the canopy of Osmanthus fragrans 'Xionghuang' on the photosynthetic characteristics
灌木种类 净光合速率(Pn)
(µmol·m−² s−1)气孔导度(Gs)
(mol·m−² s−1)胞间二氧化碳浓度(Ci)
(µmol·mol−1)蒸腾速率(Tr)
(mmol·m−² s−1)Ⅰ 2.564±0.026c 0.025±0.001b 228.726±8.139b 0.503±0.002bc Ⅱ 4.690±0.037a 0.028±0.001a 173.824±3.574c 0.638±0.039a Ⅲ 3.155±0.092ab 0.028±0.001a 175.716±3.363c 0.567±0.028b Ⅳ 3.117±0.132b 0.026±0.000b 229.232±6.233b 0.545±0.020b CK 2.417±0.040c 0.024±0.001c 289.686±29.728a 0.433±0.047c -
雄黄桂林下栽植不同灌木对雄黄桂叶片的AQY、Pnmax、Rd、LCP、LSP的影响见表4,结果表明,Ⅱ的AQY、Pnmax、LCP、LSP均为最大,而Rd为最小,且均与CK处理存在极显著差异水平(P<0.01)。对于AQY,Ⅲ极显著大于Ⅰ、Ⅳ和CK(P<0.01),Ⅰ显著大于CK(P<0.05),而Ⅳ与CK不存在显著差异。对于LCP,Ⅲ极显著大于Ⅰ、Ⅳ和CK(P<0.01),Ⅰ、Ⅳ无显著差异且均极显著大于CK(P<0.01)。对于Pnmax,Ⅲ极显著大于Ⅰ和CK(P<0.01),Ⅰ和Ⅳ均极显著大于Ⅰ和CK(P<0.01)。对于Rd,CK极显著大于Ⅲ和Ⅳ(P<0.01),而Ⅲ与Ⅳ不存在显著差异。对于LSP,Ⅲ和Ⅳ无显著差异,且均极显著大于Ⅰ和CK(P<0.01),Ⅰ极显著大于CK(P<0.01)。由此说明Ⅱ的光合作用效果最好,Ⅲ次之。
表 4 林下栽植不同灌木对雄黄桂叶片光响应特征参数
Table 4. Photosynthetic response characteristics of different shrubs planted under the canopy of Osmanthus fragrans 'Xionghuang' leaves
灌木种类 表观量子效率(AQY)
(µmol·m−2 s−1)暗呼吸速率(Rd)
(µmol·m−2 s−1)最大净光合速率(Pnmax)
(µmol·m−2 s−1)光饱和点(LSP)
(µmol·m−2 s−1)光补偿点(LCP)
(µmol·m−2 s−1)Ⅰ 0.036±0.001c 0.371±0.015a 2.733±0.155d 452.576±17.954c 5.458±0.056d Ⅱ 0.053±0.000a 0.334±0.004c 5.135±0.167a 879.636±8.580a 19.608±0.085a Ⅲ 0.041±0.001b 0.340±0.006b 3.390±0.076b 542.566±9.015b 8.818±0.154b Ⅳ 0.033±0.002d 0.365±0.007b 3.187±0.017c 528.592±3.897b 8.054±0.046c CK 0.033±0.001d 0.386±0.003a 2.444±0.044e 409.239±1.575d 3.616±0.161e -
对雄黄桂的7个枝生长量和光合指标进行Pearson相关性分析,由表5可知,枝生长量均与Ci、Rd为负相关,而与其他光合指标均呈正相关。前四次测量的枝生长量均与与Ci、Rd为极显著负相关(P<0.01)。第一次和第三次测量的枝生长量与Pn、Gs、Tr、AQY、Pnmax、LCP、LSP均为极显著正相关(P<0.01);第二次测量的枝生长量与Gs、Tr、AQY、Rd、Pnmax均为极显著正相关(P<0.01),与Pn、LCP、LSP为显著正相关(P<0.05);第四次测量的枝生长量与Pn、Tr、AQY、Rd、Pnmax、LCP、LSP均为极显著正相关(P<0.01),与Gs为显著正相关(P<0.05);第五次测量的枝生长量与Pn、Gs为显著正相关(P<0.05),与Tr、Ci的相关性为极显著水平(P<0.01),与其他光合指标无显著相关性;第六次测量的枝生长量与Pn、Gs、AQY、Rd、Ci、Rd的相关性为显著水平(P<0.05),与其他光合因子无显著相关性;第七次测量的枝生长量与7个光合指标均为无显著差异。
表 5 枝生长与光合作用的相关性
Table 5. Correlation between branch growth and photosynthesis
测量
次数净光合速率(Pn)
µmol·m−2s−1气孔导度
(Gs)
mol·m−2 s−1胞间CO2浓度(Ci)
µmol·mol−1蒸腾速率
(Tr)
mmol·m−2s−1表观量子效率(AQY)
µmol·m−2s−1暗呼吸速率(Rd)
µmol·m−2s−1最大光合速率(Pnmax)
µmol·m−2s−1光饱和点
(LSP)
µmol·m−2s−1光补偿点
(LCP)
µmol·m−2s−11 0.797** 0.825** −0.832** 0.763** 0.776** −0.918** 0.790** 0.760** 0.744** 2 0.620* 0.752** −0.741** 0.707** 0.704** −0.831** 0.659** 0.602* 0.566* 3 0.785** 0.767** −0.780** 0.722** 0.809** −0.864** 0.819** 0.773** 0.747** 4 0.898** 0.640* −0.674** 0.675** 0.890** −0.683** 0.885** 0.899** 0.881** 5 0.522* 0.525* −0.504 0.666** 0.445 −0.457 0.476 0.483 0.481 6 0.525* 0.616* −0.605* 0.514 0.602* −0.630* 0.51 0.495 0.48 7 0.412 0.343 −0.359 0.272 0.469 −0.306 0.437 0.445 0.439 注:** 在 0.01 级别,相关性极显著。* 在 0.05 级别,相关性显著。 -
植物的枝条生长是植物整体生长发育的一个重要指标,代表着植物整体生理状态和环境适应能力。以枝的生长来表征雄黄桂的整体生长,并测定雄黄桂的枝条在生长季的生长量,结果表明:林下栽植灌木对雄黄桂的生长均有促进作用,其中红枫的促进效果最明显,红花檵木次之。林下栽植不同灌木可以提高土壤微生物生物量,有助于加快土壤有机物的分解转化和养分释放[18],还可以增大凋落物自然质量,凋落物的覆盖可以维持土壤温度,增加土壤的持水率[19]。凋落物还会改变植物生长的微环境[20],当凋落物分解时,其释放出养分有助于提高土壤肥力,进而促进植物的生长。红枫促进生长的效果最好,这可能与红枫落叶的特性相关。红花檵木虽是常绿灌木,但在花朵传粉后花瓣逐渐脱落,也为植物生长提供一定的养分,而茶树和金边黄杨作为常绿灌木,凋落物量相比之下较少,促进效果不明显。
-
植物叶片的Pn、Gs、Tr等是表征光合作用的重要指标,可以反映植物的生理特性[21]。在雄黄桂林下栽植不同灌木,分析其对雄黄桂叶片的光合特性和光响应特征参数的影响。结果表明,林下栽植灌木对雄黄桂叶片的Pn、Gs、Tr、AQY、Rd、LCP和LSP均有促进作用,而Ci降低。在各项光合指标中,红枫均表现最优,而Ci极显著低于对照,这可能与红枫的落叶深根系树种特性相关,灌木的根系有助于调节土壤水分,凋落物覆盖也增加土壤的持水量[17],提供更稳定的水分供应,而在水分相对充足的情况下,植物通常能够维持较高的Gs。Gs代表着植物叶片气孔的张开程度[22],Gs增加促进了与外界的水分和气体交换,使得植物的Pn、Tr增加。随着光合速率的增加,叶片加速对CO2的消耗并将其转化为有机物,而蒸腾作用导致水分的流失,从而导致Ci降低。
-
光合作用是植物生命活动的基本过程之一,与植物的生长发育密切相关[23],是表征植物生长状况的重要指标[24]。结果表明,雄黄桂枝条的生长量与与Ci、Rd呈现负相关,而与其他光合指标均呈正相关。Gs通过调节气体交换、水分平衡和植物生长激素的运输,进而对植物的生长起到调控作用。LSP和LCP分别反映植物对强光和弱光的适应能力,AQY则是用于衡量光合作用效率的标志[25],当AQY较大时,植物中的光合色素与蛋白质复合体的数量相对较多,其对于吸收和转换光能的能力可能更为强大[26],从而提高整体光合速率;LCP越高越有利于植物在低光的生长;LSP越高则意味着植物能更高效的利用光能,产生更多的光合产物。Rd与植物枝条生长呈负相关,可能是因为Rd表示植物需要消耗能量来维持基本代谢需求,从而减少了用于植物生长发育的能量。
7个光合指标与前四次的枝生长量表现出显著或极显著差异,但从第五次开始,枝生长量与光合指标开始无显著差异,这可能是因为在生长后期,光合作用产生的有机物主要用于维持植物的代谢活动。
-
经研究发现雄黄桂林下栽植茶树、红枫、红花檵木和金边黄杨对雄黄桂的生长和光合作用都有促进作用,其中栽植红枫对雄黄桂生长的促进效果最优,其次是红花檵木,而金边黄杨和茶树的促进效果不明显。因此,在生产上解决雄黄桂生长速度相对较慢的问题时应将栽植红枫作为首选,除此之外,红枫的叶子随季节变化而呈现不同的颜色,具有较高的观赏价值;其次还可选择红花檵木,其花具有较高的观赏价值。
Effects of understory shrub on growth and photosynthesis of Osmanthus fragrans 'Xionghuang'
More Information-
摘要: 研究下层灌木(Ⅰ:茶树;Ⅱ:红枫;Ⅲ:红花檵木;Ⅳ:金边黄杨;CK:对照)对生长和光合作用的影响,确定出促进雄黄桂生长的最优灌木,为解决其生长缓慢的问题提供了科学依据。结果表明:(1)雄黄桂林下栽植以上四种灌木对其生长均有促进作用,其中Ⅱ的枝生长量高于其他灌木,Ⅲ次之,且Ⅱ第一次、第二次、第三次和第四次的枝生长量均极显著大于Ⅰ、Ⅳ和CK(P<0.01)。(2)Pn、Gs和Tr表现为Ⅱ>Ⅲ>Ⅳ>Ⅰ>CK,Ⅱ和Ⅲ的Pn和Gs均与Ⅰ、Ⅳ和CK差异达到极显著水平(P<0.01),Ⅱ的Tr显著大于Ⅰ、Ⅲ、Ⅳ(P<0.05)。(3)Ⅱ的AQY、Pnmax、LCP、LSP均为最大,而Rd为最小,其次是Ⅲ,且均与CK处理存在极显著差异水平(P<0.01)。(4)枝生长量均与Pn、Gs、Tr、AQY、Pnmax、LCP、LSP呈正相关,与Ci、Rd呈负相关。综上,在生产上解决雄黄桂生长速度相对较慢的问题时应将栽植红枫作为首选,除此之外,红枫的叶子随季节变化呈现不同的颜色,具有较高的观赏价值;其次还可选择红花檵木。Abstract: Study the understory shrub (Ⅰ: Camellia japonica; Ⅱ: Acer palmatum 'Atropurpureum'; Ⅲ: Loropetalum chinense var. rubrum; Ⅳ: Euonymus japonicus 'Aurea-marginatus'; CK: Control) influence on growth and photosynthesis, identify the best shrub to promote the growth of Osmanthus fragrans 'Xionghuang', which provides a scientific basis for solving the problem of slow growth. Results showed that (1) The above four kinds of shrubs planted under Osmanthus fragrans 'Xionghuang' had the promotio-n effect on its growth, with Ⅱ exhibiting significantly higher branch growth than other treatments. The branch growth of Ⅱ i-n the first, second, third, and fourth instances was significantly greater than Ⅰ, Ⅳ, and CK (P<0.01).(2) Pn, Gs and Tr are characterized by Ⅱ>Ⅲ>Ⅳ>Ⅰ>CK, with significant differences between Ⅱ and Ⅰ, Ⅳ, CK (P<0.01) for Pn and Gs, and Ⅱ ha-ving significantly higher Tr than Ⅰ, Ⅲ, Ⅳ (P<0.05). (3) AQY, Pnmax, LCP, and LSPwere all highest for Ⅱ, followed by Ⅲ, with significant differences from CK (P<0.01). (4) Branch growth increment was positively correlated with Pn, Gs, Tr, AQY, Pnmax, LCP, and LSP, and negatively correlated with Ci and Rd. In conclusion, planting Acer palmatum 'Atropurpureum' should be the first choice to solve the problem of relatively slow growth rate of Osmanthus fragrans 'Xionghuang' in production. In addition, the leaves of Acer palmatum 'Atropurpureum' different colors with seasonal changes, which has high ornamental value; Seco-ndly, Loropetalum chinense var. rubrum could be selected for planting.
-
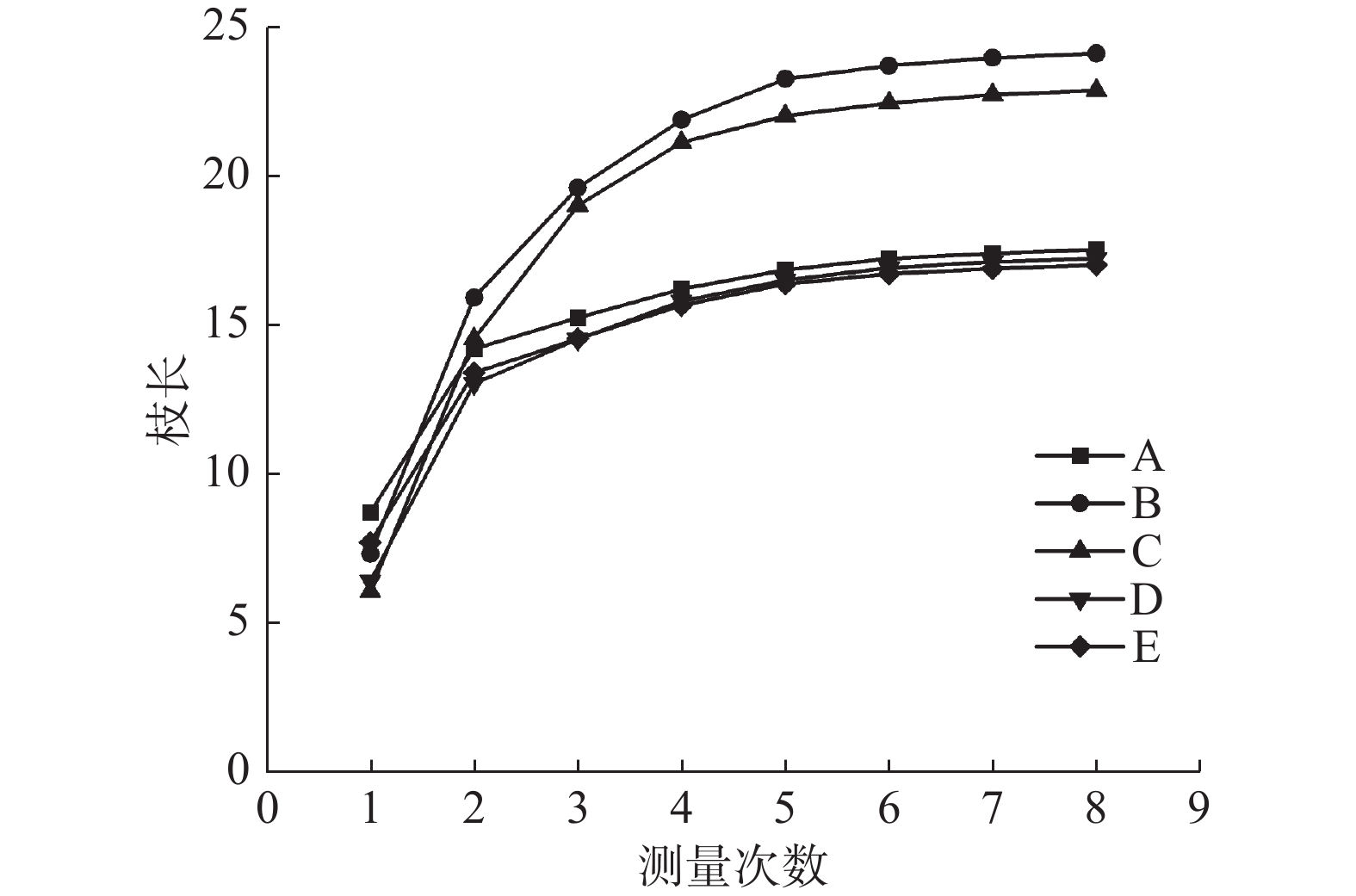
图 1 林下栽植不同灌木对雄黄桂枝生长的影响
Fig. 1 Effects of different shrubs planted under the canopy of Osmanthus fragrans 'Xionghuang' on branch growth of Osmanthus fragrans 'Xionghuang'
表 1 样地概况
Tab. 1 General situation of the sample plot
样地号 灌木种类 平均冠幅(m) 灌木平均树高(m) 平均树高(m) 平均胸径(cm) 郁闭度(%) 密度(plants·hm−2) 1-3 Ⅰ 0.523 0.960 2.34-2.66 7.73-8.59 8-13 825-875 4-6 Ⅱ 1.378 1.322 2.83-2.87 9.12-11.10 17-24 875-975 7-9 Ⅲ 0.959 1.121 2.44-2.67 7.46-8.16 12-19 850-900 10-12 Ⅳ 0.800 1.221 2.51-3.01 8.66-10.93 14-25 900 13-15 CK - - 2.38-2.75 8.45-8.69 12-13 725-875 注:Ⅰ代表茶树,Ⅱ代表红枫,Ⅲ代表红花檵木,Ⅳ代表金边黄杨,CK代表空白对照  下载: 导出CSV
下载: 导出CSV
表 2 林下栽植不同灌木对雄黄桂枝生长的影响
Tab. 2 Effects of different shrubs planted under the canopy of Osmanthus fragrans 'Xionghuang' on branch growth of Osmanthus fragrans 'Xionghuang'
灌木种类 枝生长量 1 2 3 4 5 6 7 Ⅰ 5.67±0.25c 1.14±0.06b 1.11±0.27b 0.73±0.15bc 0.38±0.06a 0.18±0.02a 0.12±0.03a Ⅱ 8.59±0.48a 3.68±0.98a 2.27±0.20a 1.37±0.19a 0.45±0.04a 0.26±0.08a 0.15±0.05a Ⅲ 8.47±0.18a 4.47±0.25a 2.11±0.11a 0.89±0.07b 0.42±0.06a 0.28±0.059a 0.13±0.04a Ⅳ 6.63±0.32b 1.47±0.17b 1.26±0.09b 0.72±0.08bc 0.41±010a 0.19±0.03a 0.12±0.02a CK 5.48±0.16c 1.04±0.20b 0.96±0.16b 0.64±0.09c 0.33±0.07a 0.18±0.0a 0.12±0.02a
下载: 导出CSV
表 3 林下栽植不同灌木对雄黄桂光合特性的影响
Tab. 3 Effects of different shrubs planted under the canopy of Osmanthus fragrans 'Xionghuang' on the photosynthetic characteristics
灌木种类 净光合速率(Pn)
(µmol·m−² s−1)气孔导度(Gs)
(mol·m−² s−1)胞间二氧化碳浓度(Ci)
(µmol·mol−1)蒸腾速率(Tr)
(mmol·m−² s−1)Ⅰ 2.564±0.026c 0.025±0.001b 228.726±8.139b 0.503±0.002bc Ⅱ 4.690±0.037a 0.028±0.001a 173.824±3.574c 0.638±0.039a Ⅲ 3.155±0.092ab 0.028±0.001a 175.716±3.363c 0.567±0.028b Ⅳ 3.117±0.132b 0.026±0.000b 229.232±6.233b 0.545±0.020b CK 2.417±0.040c 0.024±0.001c 289.686±29.728a 0.433±0.047c
下载: 导出CSV
表 4 林下栽植不同灌木对雄黄桂叶片光响应特征参数
Tab. 4 Photosynthetic response characteristics of different shrubs planted under the canopy of Osmanthus fragrans 'Xionghuang' leaves
灌木种类 表观量子效率(AQY)
(µmol·m−2 s−1)暗呼吸速率(Rd)
(µmol·m−2 s−1)最大净光合速率(Pnmax)
(µmol·m−2 s−1)光饱和点(LSP)
(µmol·m−2 s−1)光补偿点(LCP)
(µmol·m−2 s−1)Ⅰ 0.036±0.001c 0.371±0.015a 2.733±0.155d 452.576±17.954c 5.458±0.056d Ⅱ 0.053±0.000a 0.334±0.004c 5.135±0.167a 879.636±8.580a 19.608±0.085a Ⅲ 0.041±0.001b 0.340±0.006b 3.390±0.076b 542.566±9.015b 8.818±0.154b Ⅳ 0.033±0.002d 0.365±0.007b 3.187±0.017c 528.592±3.897b 8.054±0.046c CK 0.033±0.001d 0.386±0.003a 2.444±0.044e 409.239±1.575d 3.616±0.161e
下载: 导出CSV
表 5 枝生长与光合作用的相关性
Tab. 5 Correlation between branch growth and photosynthesis
测量
次数净光合速率(Pn)
µmol·m−2s−1气孔导度
(Gs)
mol·m−2 s−1胞间CO2浓度(Ci)
µmol·mol−1蒸腾速率
(Tr)
mmol·m−2s−1表观量子效率(AQY)
µmol·m−2s−1暗呼吸速率(Rd)
µmol·m−2s−1最大光合速率(Pnmax)
µmol·m−2s−1光饱和点
(LSP)
µmol·m−2s−1光补偿点
(LCP)
µmol·m−2s−11 0.797** 0.825** −0.832** 0.763** 0.776** −0.918** 0.790** 0.760** 0.744** 2 0.620* 0.752** −0.741** 0.707** 0.704** −0.831** 0.659** 0.602* 0.566* 3 0.785** 0.767** −0.780** 0.722** 0.809** −0.864** 0.819** 0.773** 0.747** 4 0.898** 0.640* −0.674** 0.675** 0.890** −0.683** 0.885** 0.899** 0.881** 5 0.522* 0.525* −0.504 0.666** 0.445 −0.457 0.476 0.483 0.481 6 0.525* 0.616* −0.605* 0.514 0.602* −0.630* 0.51 0.495 0.48 7 0.412 0.343 −0.359 0.272 0.469 −0.306 0.437 0.445 0.439 注:** 在 0.01 级别,相关性极显著。* 在 0.05 级别,相关性显著。
下载: 导出CSV
-
[1] 徐珊珊. 观赏园艺植物在城市绿化中的应用[J]. 智慧农业导刊,2022,2(4):36−38. [2] 贺维,杨育林,高鹏,等. 岷江干旱河谷两种主要人工林林下豆科物种组成及植被多样性[J]. 四川林业科技,2022,43(5):41−47. He W, Yang Y L, Gao P, et al. Species composition and vegetation diversity of Leguminosae species under two main plantations in arid valley of the Minjiang river[J]. Journal of Sichuan Forestry Science and Technology, 2022, 43(5): 41−47. [3] Giuggiola A, Zweifel R, Feichtinger L M, et al. Competition for water in a xeric forest ecosystem-Effects of understory removal on soil micro-climate, growth and physiology of dominant Scots pine trees[J]. Forest Ecology and Management, 2018, 409: 241−249. doi: 10.1016/j.foreco.2017.11.002 [4] Liao Y C, Fan H B, Wei X H, et al. Competition increased fine root biomass in Chinese fir (Cunninghamia lanceolata) plantations in Subtropical China[J]. Forest Ecology and Management, 2019, 435: 151−157. doi: 10.1016/j.foreco.2018.12.035 [5] Messier C, Coll L, Poitras-Larivière A, et al. Resource and non-resource root competition effects of grasses on early-versus late-successional trees[J]. Journal of Ecology, 2009, 97(3): 548−554. doi: 10.1111/j.1365-2745.2009.01500.x [6] 李钢,梁音,曹龙熹. 次生马尾松林下植被恢复措施的水土保持效益[J]. 中国水土保持科学,2012,10(6):25−31. doi: 10.3969/j.issn.1672-3007.2012.06.005 Li G, Liang Y, Cao L X. Effects of different vegetation restoration patterns on soil erosion in secondary Pinus massoniana pure forest[J]. Science of Soil and Water Conservation, 2012, 10(6): 25−31. doi: 10.3969/j.issn.1672-3007.2012.06.005 [7] 施秀珍,王建青,黄志群,等. 树种多样性对土壤微生物群落结构和元素生物地球化学循环的影响研究进展[J]. 生态学报,2022,42(15):6092−6102. Shi X Z, Wang J Q, Huang Z Q, et al. Research progresses in the effects of tree species diversity on soil microbialcommunities and biogeochemical cycling of elements[J]. Acta Ecologica Sinica, 2022, 42(15): 6092−6102. [8] 郭琦,王新杰. 不同混交模式杉木人工林林下植被生物量与土壤物理性质研究[J]. 中南林业科技大学学报,2014,34(05):70−74. Guo Q, Wang X J. Undergrowth biomass and soil physical properties under Cunninghamia lanceolata with different patterns of mixed forests[J]. Journal of Central South University of Forestry & Technology, 2014, 34(05): 70−74. [9] 苟蓉,游明鸿,白史且,等. 雌雄构树幼苗光合能力对干旱胁迫响应的性别差异[J]. 四川林业科技,2020,41(6):1−7. Gou R, You M H, Bai S Q, et al. Gender differences on photosynthetic capacity of Broussonetia papyrifera seedlings in response to drought stress[J]. Journal of Sichuan Forestry Science and Techznology, 2020, 41(6): 1−7. [10] 董立格,王贤荣,丁雨龙. 桂花花期物候研究[J]. 南京林业大学学报自然科学版,2014,38(S1):51−56. Dong L G, Wang X R, Ding Y L. Study on the Osmanthus fragrans blooming season phenology[J]. College of Biology and the Environment, Nanjing Forestry University, Nanjing, 2014, 38(S1): 51−56. [11] 唐新瑶,亢亚超,梁喜献,等. 氮磷钾配比施肥对观光木幼苗生理与光合特性的影响[J]. 西北林学院学报,2022,37(4):37−42. Tang X Y, Kang Y C, Liang X X, et al. Effects of N, P and K Proportional Fertilization on the Physiological and Photosynthetic Characteristics of Tsoongiodendron odorum Seedlings[J]. JOURNAL OF NORTHWEST FORESTRY UNIVERSITY, 2022, 37(4): 37−42. [12] 闫小莉,王德炉. 不同类型土壤栽培对苦丁茶树叶片生长和光合特性的影响[J]. 生态学报,2019,39(19):7208−7217. Yan X L, Wang D L. Effects of different soil types on growth and photosynthetic characteristics of <italic> Ligustrum robustum</italic>[J]. Acta Ecologica Sinica, 2019, 39(19): 7208−7217. [13] 田秋玲,乐佳兴,吴焦焦,等. 西南丘陵地区紫色土酸性对无患子幼树生长和光合特性的影响[J]. 生态学报,2020,40(11):3756−3763. Tian Q L, Yue J X, Wu J J, et al. Effects of southwest hilly areas’s purple soil acidity on the growth and photosynthetic characteristics of <italic> Sapindus mukorossi</italic> Gaertn saplings[J]. Acta Ecologica Sinica, 2020, 40(11): 3756−3763. [14] 梁俊林,毛绘友,郭丽,等. 遮阴对3种珍贵乡土阔叶树种幼苗生长及光合作用的影响[J]. 西北林学院学报,2019,34(4):57−63. Liang J L, Mao H Y, Guo L, et al. Influence of Shading on the Seedling Growth of Three Precious Native Tree Species and Photosynthesis[J]. JOURNAL OF NORTHWEST FORESYTY UNIVERSITY, 2019, 34(4): 57−63. [15] 陈斌,刘筱玮,贾琳,等. 光强对4种鸭跖草科植物生长和光合特性的影响[J]. 生态学报,2022,42 (4):1450-1461. Chen B, Liu X W, Jia L, et al. Effects of light intensity on the growth and photosynthetic characteristics of four Commelinaceae plants. Acta Ecologica Sinica, 2022, 42(4): 1450-1461. [16] 方精云,王襄平,沈泽昊,等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性,2009(6):533−548. Jing Y F, Xiang P W, Ze H S, et al. Methods and protocols for plant community inventory[J]. Biodiversity Science, 2009(6): 533−548. [17] Ye Z P. A new model for relationship between light intensity and the rate of photosynthesis in Oryza sativa[J]. Photosynthetica, 2007, 45(4): 637−640. doi: 10.1007/s11099-007-0110-5 [18] 张贾宇,佘婷,鄂晓伟,等. 杨树人工林幼林阶段林下植被管理对土壤微生物生物量碳、氮酶活性的影响[J]. 生态学报,2021,41(24):9898-9909. Zhang J Y, She T, E X W, et al. Effects of understory vegetation management on soil microbial biomass carbon and nitrogen and extracellular enzyme activities in the early stages of poplar plantation. Acta Ecologica Sinica, 2021, 41(24): 9898-9909. [19] 赵冲,蔡一冰,黄晓,等. 杉木凋落物覆盖对自身幼苗出土和早期生长的影响[J]. 应用生态学报,2019,30(2):481−488. Zhao C, Cai Y B, Huang X, et al. Effects of Chinese fir litter cover on its seedling emergence and early growth[J]. Chinese Journal Of Applied Ecology, 2019, 30(2): 481−488. [20] Asplund J, Hustoft E, Nybakken L, et al. Litter impair spruce seedling emergence in beech forests: a litter manipulation experiment[J]. Scandinavian Journal of Forest Research, 2018, 33(4): 332−337. doi: 10.1080/02827581.2017.1388440 [21] 付忠,谢世清,徐文果. 等. 不同光照强度下谢君魔芋的光合作用及能量分配特征[J]. 应用生态学报,2016,27(4):1177−1188. Fu Z, Xie S Q, Xu W G, et al. Characteristics of photosynthesis and light energy partitioning in Amorphophallus xiei grown along a light-intensity gradient[J]. Scandinavian Journal of Forest Research, 2016, 27(4): 1177−1188. [22] 刘玉娟. 盐胁迫对柴达木盆地几种灌木的生理影响[D]. 北京林业大学,2015. [23] 戴建昊,韩暖,余飞燕,等. 不同株行距对核桃光合作用的影响[J]. 西北林学院学报,2019,34(4):96−100. doi: 10.3969/j.issn.1001-7461.2019.04.14 Dai J H, Han N, Yu F Y, et al. Effects of Different Row Spacing on Photosynthesis of Walnut Trees[J]. JOURNAL OF NORTHWEST FORESYTY UNIVERSITY, 2019, 34(4): 96−100. doi: 10.3969/j.issn.1001-7461.2019.04.14 [24] 宋淑珍,熊康宁,池永宽,等. 喀斯特石漠化地区菊苣光合特性年际变化研究[J]. 中国草地学报,2017,39(02):65−70. [25] 惠俊爱,党志,叶庆生. 镉胁迫对玉米光合特性的影响[J]. 农业环境科学学报,2010,29(2):205−210. Hui J A, Dang Z, Ye Q S. Influence of Cadmium Stress on Photosynthetic Characteristics of Maize[J]. JOURNAL OF AGRO-ENVIRONMENT SCIENCE, 2010, 29(2): 205−210. [26] Richardson A D, Berlyn G P. Spectral reflectance and photosynthetic properties of <italic> Betula papyrifera</italic> (Betulaceae) leaves along an elevational gradient on Mt. Mansfield, Vermont, USA[J]. American Journal of Botany, 2002, 89(1): 88−94. doi: 10.3732/ajb.89.1.88 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 19
- HTML全文浏览量: 4
- PDF下载量: 2
- 被引次数: 0









