






 下载:
下载:

-
栖息地是物种生存栖息的空间,是能够为物种提供食物、庇护和繁殖机会的场所,当前保护物种最有效的办法之一就是保护它们赖以生存的栖息地[1]。日活动节律指动物在一天中的活动强度随时间发生的周期性变化[2]。栖息地和日活动节律是物种长期适应环境的结果,主要受到物种本身的食物需求以及生物钟等特性的影响,也因气候、捕食和竞争关系等外部环境的改变产生波动[3-4]。探究物种栖息地选择模式和活动节律的变化,不仅能够了解物种本身的生物学特性,还能够有助于分析物种之间的相互关系,这对物种乃至整个生态系统的保护都有十分重要的意义。
物种分布模型(SDM)是可以把物种地理分布与环境因子相联系的一种重要工具,近年来被广泛应用在物种潜在栖息地研究当中[5-7],在SDM中当前最适用的是最大熵模型(MaxEnt)[6][8-9],其通过环境变量数据以及目标物种已有的分布数据,对目标物种进行潜在栖息地分析预测,最终根据最大熵理论的原理,推测出该目标物种的潜在适宜栖息地[10-14]。
近年来, 在国内外大中型兽类和鸟类监测研究中,红外相机技术已得到普遍应用,红外相机后期数据分析中常使用相对多度指数(RAI)来评估某物种的相对种群数量[15-16],RAI =(独立探测次数/相机总有效工作天数)×1,000。单个位点上拍摄到某物种记为该物种的一次有效探测,30 min内同一位点拍摄到的相同物种算作同一次有效探测。红外相机监测累积了大量含有时间信息的野生动物行为原始数据, 极大地促进了在野生动物活动节律方面的深入探讨[17-19]。目前, 国内外利用红外相机数据开展物种活动节律研究主要采取核密度估计方法(kernel density estimation), 主要用于描述单物种以及多物种的活动节律和重叠程度[20-23],以陆生哺乳动物和地栖性鸟类等类群为主[24]。
红腹角雉(Trgopan temminckii)属鸡形目(Galliformes)雉科(Phasianidae)鸟类,是主要分布于我国的国家Ⅱ级重点保护野生雉类[25]。从20世纪80年代开始,科研人员陆续发表红腹角雉研究相关论文,主要涉及种群结构[26-28]、食性分析[29]、生境选择[30-31] 、繁殖习性[27][32]、活动节律[33-34]等方面,但对野外红腹角雉个体栖息地选择和季节活动节律差异的研究资料较为缺乏。
关于卧龙国家级自然保护区红腹角雉的研究,仅见张福成等[35]、李仁贵等[36]对卧龙“五一棚”附近区域红腹角雉与其他鸟类的种间关系和红腹角雉冬季栖息生境选择进行了研究。本文收集了卧龙国家级自然保护区2014—2019年红外相机调查中采集到的红腹角雉相关信息,研究保护区内野生红腹角雉的潜在适宜栖息地和活动节律差异,以期为保护区红腹角雉乃至同域分布的其他雉类的保护与管理提供科学依据。
-
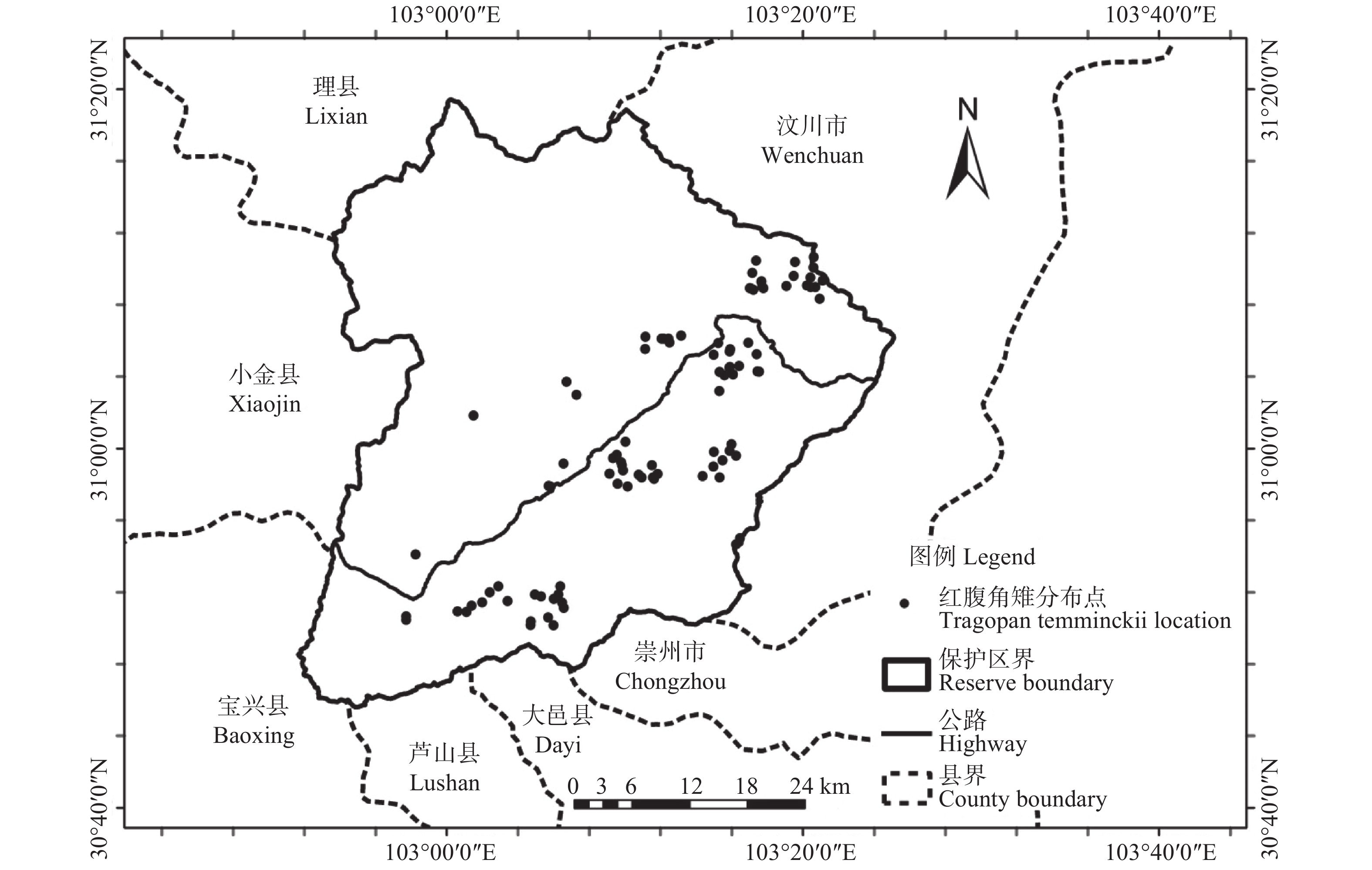
四川卧龙国家级自然保护区位于北纬30°45′—31°25′,东经102°51′—103°24′之间,东西长52 km、南北宽62 km,全区总面积2 000 km2,地处邛崃山脉东南坡,属四川盆地向川西高原过渡地带,地形复杂,海拔跨度大,植被类型丰富,是我国建立最早的以保护大熊猫(Ailuropoda melanoleuca)等珍稀野生动物及高山森林生态系统为主的综合性自然保护区[37]。根据保护区近年监测研究,共记录到红腹角雉、绿尾虹雉(Lophophorus lhuysii)、红腹锦鸡(Chrysolophus pictus)、勺鸡(Pucrasia macrolopha)、血雉(Ithaginis cruentus)、红喉雉鹑(Tetraophasis obscurus)、雪鹑(Lerwa lerwa)等鸡形目雉类12种。红腹角雉为保护区种群数量较大,最为常见的雉类之一,实体遇见率较高,常于中低海拔针阔混交林和阔叶林中灌木盖度低、竹林少的区域发现其活动痕迹。
-
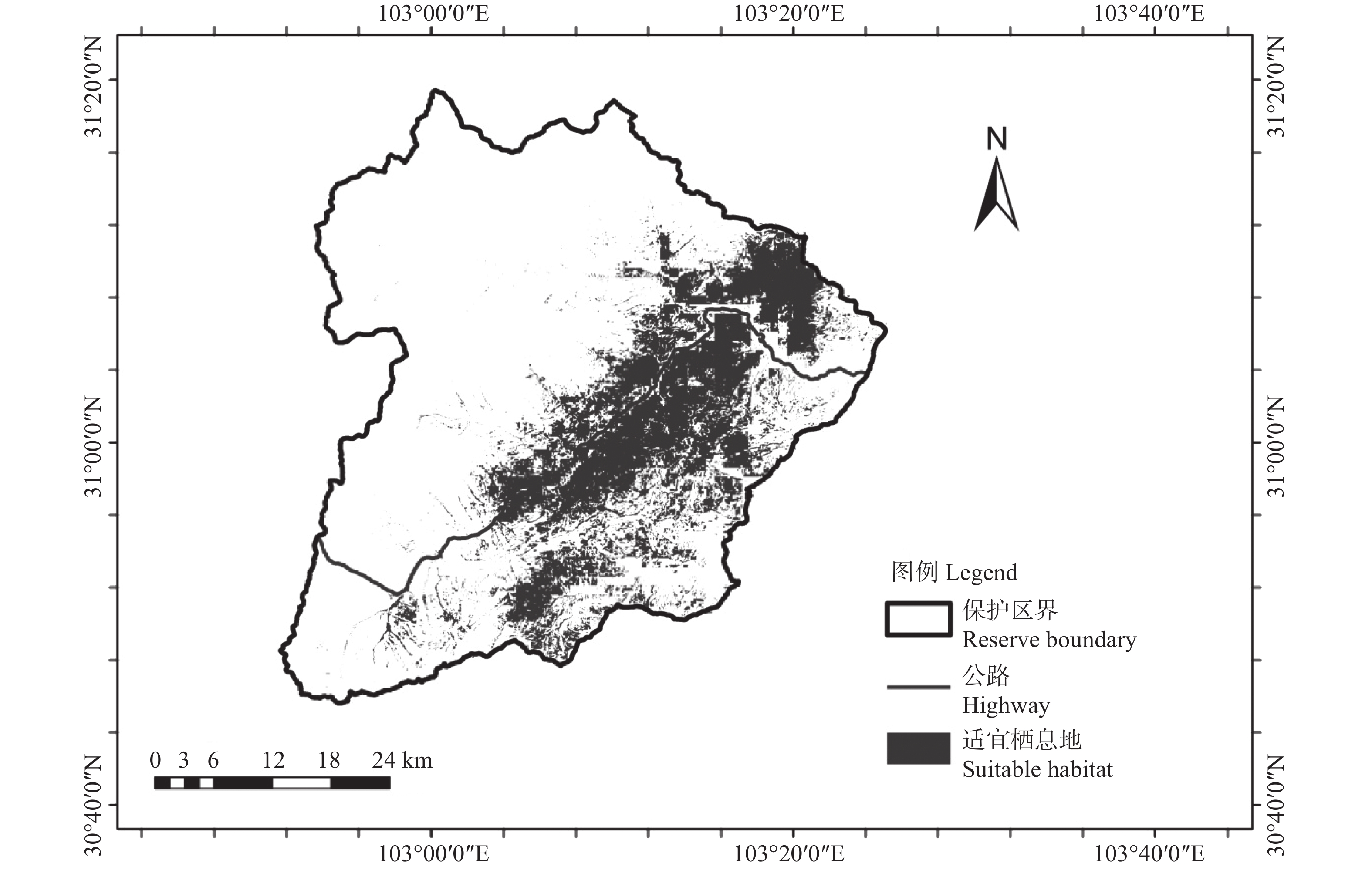
收集了卧龙保护区2014年至2019年全区范围野外布设红外相机数据。据统计,共503个有效相机调查位点,覆盖海拔范围1 470~4 780 m,相机总有效工作日126 286 d,共获得兽类、鸟类的照片和视频269 979份,有效探测数37 779次。共鉴定出分属6目17科的38种野生兽类和分属7目22科的77种野生鸟类。503个有效位点中,共收集红腹角雉红外相机记录位点100个(见图1),覆盖海拔范围1 802~4 254 m,共计拍摄红腹角雉照片和视频2686份,有效探测数549次。

图 1 研究区域地理位置及红腹角雉记录位点示意图
Figure 1. Schematic map of the study area and the occurrence locations of Trgopan temminckii in the study area
-
利用MaxEnt模型(版本3.4.0)模拟红腹角雉的栖息地分布,选取气候、地形、植被、人类影响四大类因子作为模型预测的环境变量。气候数据来源于WorldClim网站的Bioclim数据集中的19个气候图层,分辨率为30arc-seconds。在SPSS软件中对研究区域内这19个气候变量的person相关性进行检验,去除相关系数高于0.7的变量,以减小变量共线性对模型结果的影响[38]。地形数据来源于中国科学院地理空间数据云平台的数字高程(DEM)图,分辨率为30 m×30 m。从获取的数字高程图中,利用ArcMAP 10.4的三维分析工具得到相应的坡向、坡度变量图层,利用空间分析工具对坡度、坡向进行重分类(见表1)。植被数据来源于Land Processes Distributed Active Archive Center的MODIS数据产品MCD12Q1和MOD13Q1 。从MCD12Q1数据集提取分辨率为500 m×500 m的植被类型数据,在ArcMAP中对植被类型进行重分类(见表1));从MOD13Q1数据集提取分辨率为250 m×250 m的植被指数NDVI (归一化植被指数)。人类干扰衡量指标采用人口密度指数,数据来源为中科院资源环境数据云平台,原始分辨率为1 km×1 km。所有环境变量图层的分辨率采用最近邻法统一重采样为30 m×30 m,并转为ASC格式,用于后续的建模分析。在本研究中,因为动物位点在研究区域内分布较为分散,且分布数据量有限,未对分布点做疏化处理,全部用于后续的分布模型。在模型中设置训练数据的比例为75%,其余25%为检验数据,重复运行10次Bootstrap,选择创建响应曲线(Response curves),并对变量重要性做刀切分析(Jackknife)。在建模过程中,首先把所有选定变量放入模型进行预分析,选取对预测结果贡献较大的因子,最终保留海拔、坡度、坡向、bio2(月均昼夜温)、bio11(最冷季度平均温度)、bio13(最湿月降水量)、bio14(最干月降水量)、bio18(最暖季度降水量)、人口密度、NDVI指数和植被类型共11个变量(见表1),使用预筛后的11个变量对研究区域内的红腹角雉的空间分布构建最终模型。
表 1 红腹角雉适宜栖息地预测最终模型中使用的环境变量
Table 1. Environmental variables used in the final model of suitable habitat prediction of Trgopan temminckii
环境变量
Environmental variables分类与赋值
Classification and assignment原始分辨率
Original resolution来源
Source地形Topography 海拔 Elevation 连续变量Continuous variable 30 m 中国科学院计算机网络信息中心地理空间数据云平台Geospatial Data Gloud 坡度 Slope 0°–10° (1); 10°–20° (2); 20°–30° (3);30°–45° (4); > 45° (5) 坡向 Aspect 45°–225°(1) ; 225°–45°(2) 气候Climate 月均昼夜温Mean diurnal range (bio2) 连续变量Continuous variable 30″ WorldClim 最冷季度平均温度 Mean Temperature of Coldest Quarter (bio11) 连续变量Continuous variable 最湿月降水量 Precipitation of Wettest Month(bio13) 连续变量Continuous variable 最干月降水量Precipitation of Driest Monthbio (14) 连续变量Continuous variable 最暖季度降水量Precipitation of Warmest Quarter (bio18) 连续变量Continuous variable 植被Vegetation 归一化植被指数Normalized differenceVegetation index (NDVI) 连续变量Continuous variable 250 m Land Processes Distributed Active Archive Center 植被类型Land cover 高山流石滩 Alpine screes (1)
草甸草地 Meadow grassland (2)
高山灌丛 Alpine shrub (3)
针叶林 Coniferous forest (4)
针阔混交林Coniferous and broad-leaved mixed forest (5)
阔叶林 Broad-leaved forest (6)
其他 Others (7)500 m 人类干扰anthropogenic disturbance 人口密度Population density 连续变量Continuous variable 1 km 中科院资源环境数据云平台Resource and environment Data Gloud 以模型结果中给定的特异度和敏感度的最大和(Specificity logistic threshold plus maximum training sensitivity)作为阈值[39],在ArcMap10.4中将红腹角雉的预测结果分别重分类为适宜栖息地与非适宜栖息地,适宜栖息地为高于该物种分布阈值的区域,非适宜栖息地为低于该阈值的区域。
以ROC曲线(Receiver operating charac-teristic curve)中的AUC值(Area under curve)来评价模型的模拟成效。0.5~0.6为不合格,0.6~0.7为较差,0.7~0.8为一般,0.8~0.9为较好,0.9~1.0为优秀。
-
根据保护区的物候特点,分别定义3—5月、6—8月、9—11月、12月至来年2月为春季、夏季、秋季、冬季。特定时间段内动物被红外相机捕捉到的几率与动物活动强度成正比[40],因此可以用RAI数值大小来反映红腹角雉某一个时间段的活动强度。
日活动节律的数据类型是以24h为一周期,研究中主要采用非参数核密度估计方法对该类数据进行分析,对连续时间分布的动物行为事件随机取样,核密度估计方法是从无任何假定的原始数据本身出发研究数据分布规律的方法[20]。采用条件循环核密度方法绘制差异化的等值曲线来表述各时间段动物活动强度[21]。采用核密度估计方法计算不同季节活动节律的重叠系数。重叠系数是通过对比至少2个周期长度的密度函数的各时间点而得到[20]。参照Schmid和Schmidt[41]提出的方法,根据样本量大小,计算不同季节间红腹角雉日活动节律重叠度Δ。在R软件中使用overlap包绘制红腹角雉在不同季节的日活动节律曲线,并计算不同季节间重叠度,使用activity包计算红腹角雉在不同季节的日活动节律曲线差异性(P值)。
-
本研究平均AUC值为0.883,表明 MaxEnt 模型预测红腹角雉栖息地潜在分布具有较高的精度。预测得到红腹角雉适宜栖息地面积为430.38 km2,主要分布在保护区公路(皮条河)两侧、中河流域、西河流域,占保护区总面积的21.52%(见图2)。从影响红腹角雉的环境变量上看,11个变量中,对红腹角雉分布影响最大的因子为植被类型、 bio2和坡度,其贡献率依次为41.4%、21.5%和11.4%。红腹角雉偏好活动的区域海拔为2 000~3 000 m,在7种植被类型中最为偏好针阔混交林,其次为阔叶林和针叶林;坡度中,红腹角雉偏好活动于坡度小,较为平缓的区域,分布概率随坡度的增加逐渐减小;在月均昼夜温差(bio2)为10 ℃时,红腹角雉分布概率最大。
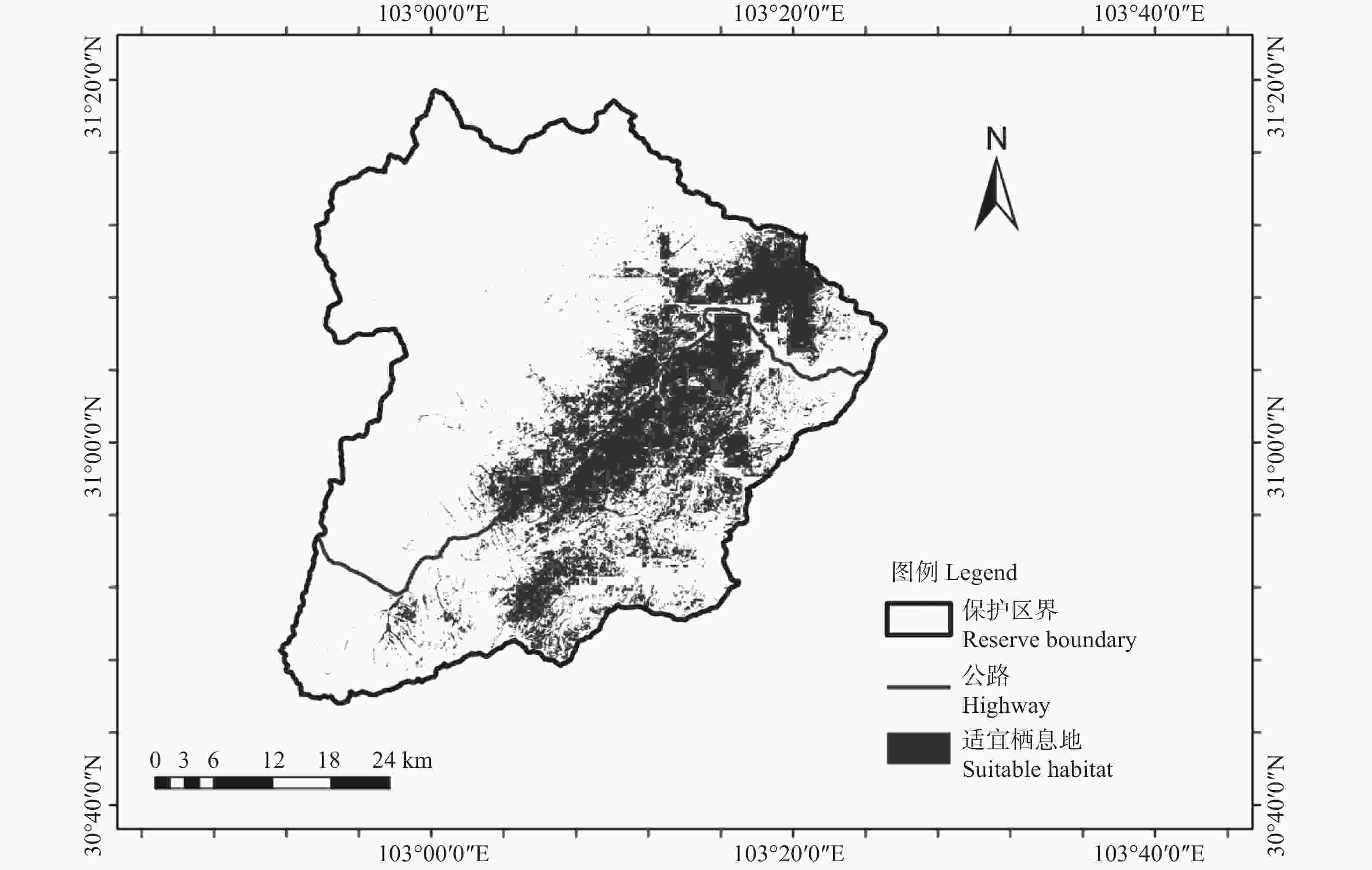
图 2 红腹角雉栖息地分布
Figure 2. Habitat distribution of Trgopan temminckii
-
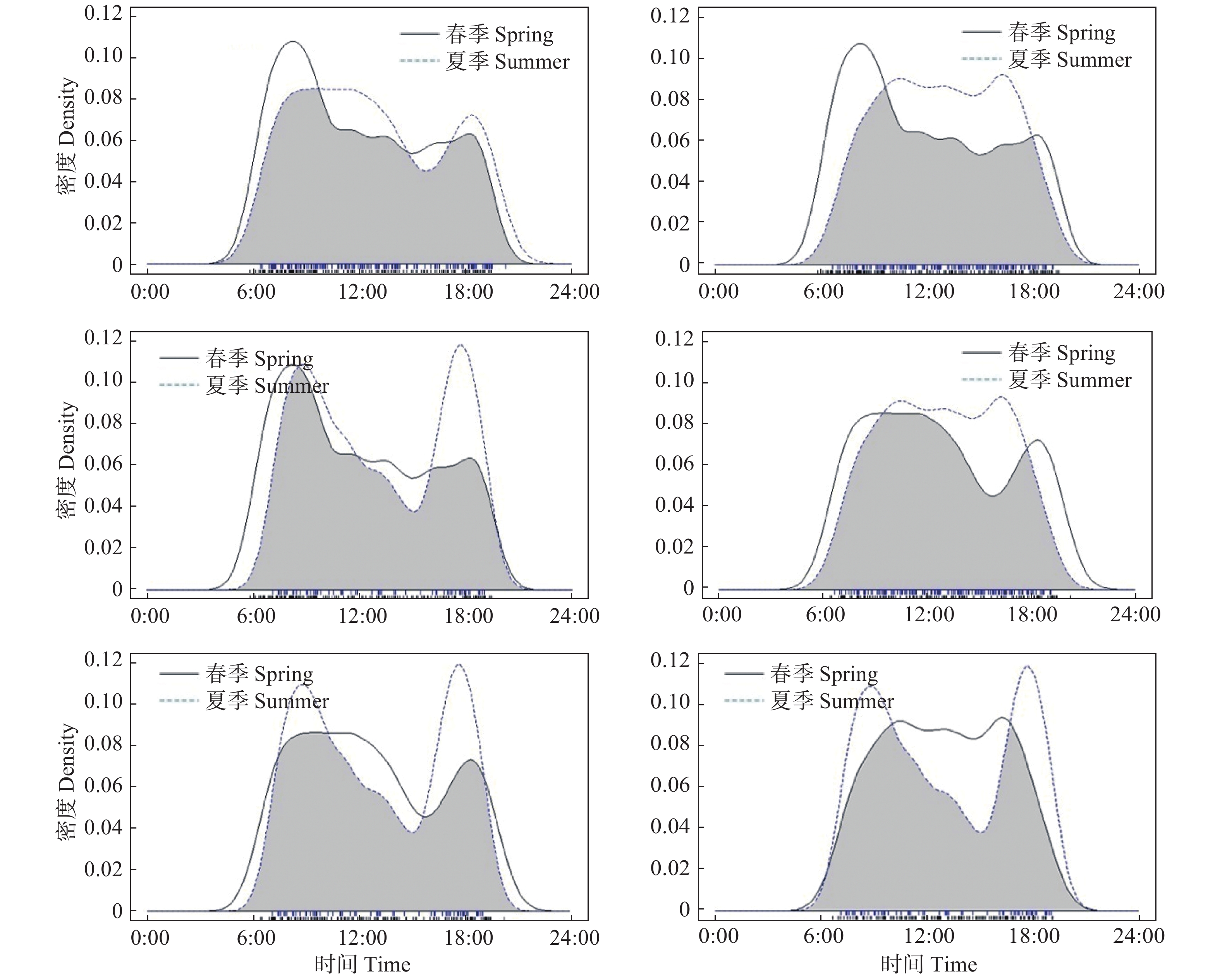
红腹角雉的549 次有效探测数中,春季有效探测数为188次,夏季有效探测数为142次,秋季有效探测数为153次,冬季有效探测数为66次,其RAI指数分别为1.49、1.12、1.21、0.52。说明红腹角雉在春季活动强度最高,秋季、夏季次之,冬季最低。日活动节律分析表明:红腹角雉在春夏秋冬四季均表现为明显的昼行性,春季在8:00和18:00 出现2个活动高峰;夏季在8:00—12:00和18:00出现2个活动高峰;秋季活动高峰在10:00—17:00;冬季在9:00和18:00出现2个活动高峰(见图3)。
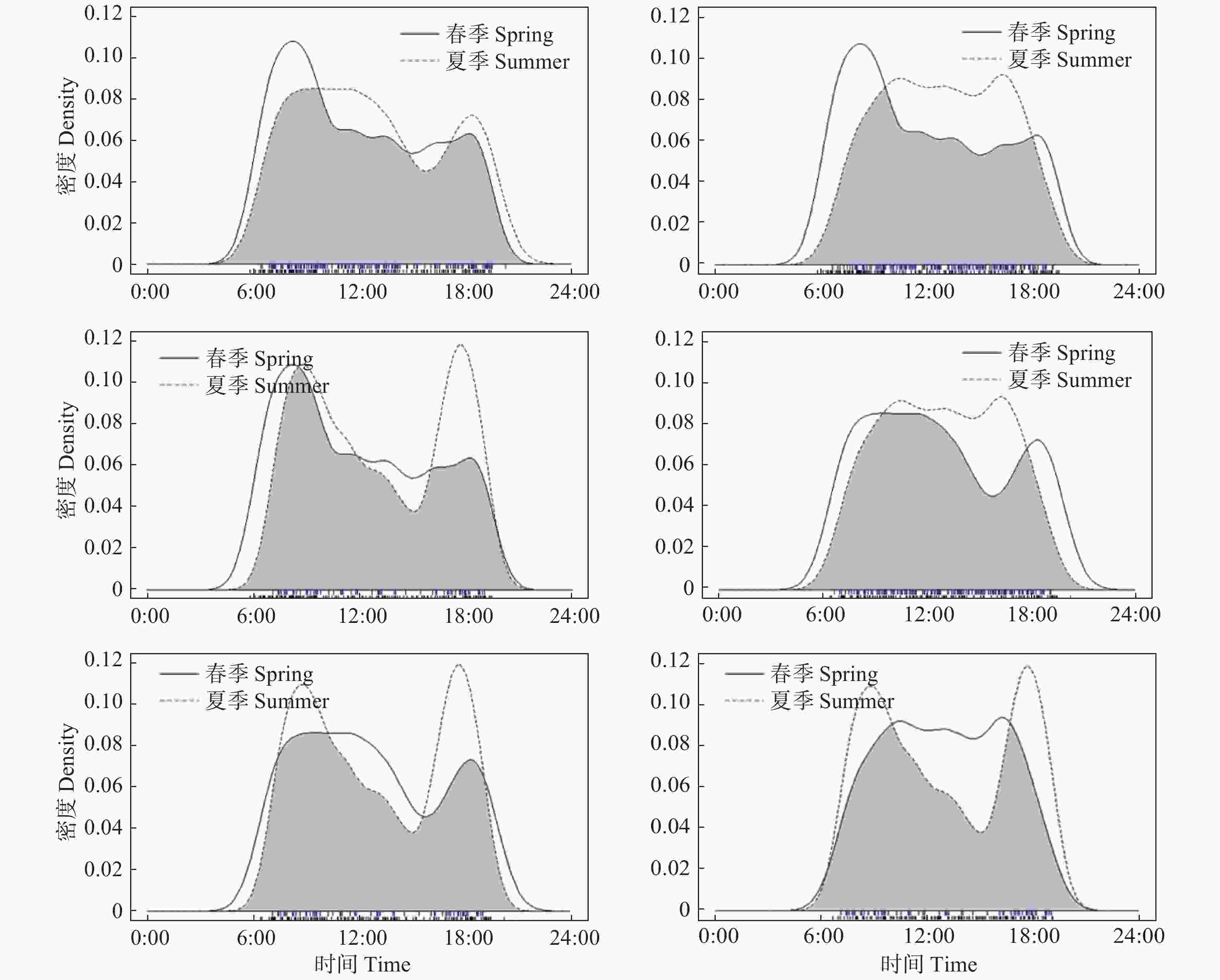
图 3 红腹角雉在春季、夏季、秋季、冬季的活动节律曲线比较( 灰色为重叠区域)
Figure 3. Comparison of daily activity rhythm of Trgopan temminckii in spring, summer, autumn and winter (The overlapping coefficient equals the area in grey)
在对不同季节间红腹角雉的日活动节律重叠度分析中选择Δ4作为估计参数,差异性检测中使用P值作为衡量指标。结果表明,春季与夏季日活动节律重叠度估计值为0.89(重叠度取值为0~1,值越大,日活动节律重叠度越高),P值为0.22,日活动节律无明显差异;春季与秋季日活动节律重叠度估计值为0.80,P值小于0.01,日活动节律差异明显;春季与冬季日活动节律重叠度估计值为0.85,P值为0.18,日活动节律无明显差异;夏季与秋季日活动节律重叠度估计值为0.85,P值为0.02,日活动节律差异明显;夏季与冬季日活动节律重叠度估计值为0.86,P值为0.19,日活动节律无明显差异;秋季与冬季日活动节律重叠度估计值为0.80,P值为0.01,日活动节律差异明显(见图3)。以上结果表明,红腹角雉在秋季的日活动节律与春季、夏季、冬季差异明显,而春季、夏季、冬季之间红腹角雉活动节律无明显差异。
-
预测得到卧龙国家级自然保护区红腹角雉潜在适宜栖息地总面积430.38 km2,占保护区总面积的21.52%,说明红腹角雉在保护区分布较为广泛,主要分布在保护区皮条河两侧、中河、西河中低海拔区域。环境因子中对红腹角雉栖息地适宜度贡献最大的为植被类型,红腹角雉最偏好的栖息地为针阔混交林,其次为阔叶林和针叶林,偏好活动的区域海拔为2 000~3 000 m,正好也是保护区针阔混交林和阔叶林分布的海拔范围。保护区针阔混交林、阔叶林、针叶林分布区域植被丰富度高于高海拔的灌丛、草地,且温度较适宜,方便红腹角雉取食,同时离人为活动区域较远,干扰较小。卧龙保护区红腹角雉偏好坡度较平缓的区域活动,分析原因可能为平缓区域掉落的枝叶、果实等食物相对较多且方便行走,更容易获取食物。
活动节律是动物对外界自然环境变化采取的适应性对策,也是动物不断进化的体现[42]。随着季节的更迭,多数野生动物的活动强度会随着发情、求偶、交配、产仔、育幼、越冬等行为活动的产生而发生周期变化。红腹角雉活动强度在春季最高,秋季、夏季次之,冬季最低,这可能与红腹角雉的生活史以及保护区内食物丰富度有关。一是,红腹角雉于每年4月份开始寻觅交配对象,求偶,5—6 月产卵、孵化,7—9 月育幼[32],此段时间活动强度较大,易被红外相机拍摄到;另一个,红腹角雉以取食植物为主[29],4—10月是卧龙保护区内内食物最丰盛的月份,尤其是红腹角雉喜食的植物嫩芽、枝、果实等都集中出现。在红腹角雉栖息地模型预测中,月均昼夜温差10 ℃时其分布概率最大,根据卧龙气候特征,红腹角雉适宜生境在春秋两季较大可能会出现10 ℃左右温差,符合红腹角雉活动强度变化规律。此外,冬季RAI指数相对偏低,可能是由于卧龙保护区冬季中高海拔段气温较低且有积雪覆盖,红腹角雉觅食困难,此时的红腹角雉多向未有积雪覆盖的低海拔区域迁徙,而本次研究中,低海拔红外相机布设数量相对较少,可能造成冬季无法捕捉到更多红腹角雉红外相机影像。建议后续研究增加红腹角雉低海拔红外相机影像数据量收集,进一步分析论证红腹角雉季节性海拔迁移规律。
红腹角雉季节活动节律差异分析采用的核密度估计方法参照陈立军等[23]中3种鸡形目鸟类的日活动节律及其重叠程度分析方法。结果表明,红腹角雉在春夏秋冬四季日活动节律重叠度均较高,表现明显的昼行性,很少拍到红腹角雉夜间活动情况,一个是与红腹角雉夜晚主要栖息于树上且多静止不动的行为有关[30],另一个,红外相机多安放于离地面不高处,难以拍摄到树栖动物。红腹角雉在秋季的日活动节律与在春季、夏季、冬季的日活动节律,均表现出显著差异。春季、夏季、冬季的红腹角雉在上午各有1个活动峰期,可能与其觅食规律相关,即这个时段可能是上午觅食高峰期,之后阳光照射不断增强,温度升高,红腹角雉活动强度降低,进入一段休息时间,下午会再出现1个小活动高峰,可能为夜间树栖补充能量有关。秋季红腹角雉活动高峰一直从上午10:00持续到下午17:00,分析原因可能为秋季是卧龙保护区果实成熟、枝叶脱落的季节,食物资源丰富,且此时天气较好,温度湿度适宜,适合较长时间活动觅食,同时也可能是为越冬尽可能多的储备脂肪。
利用物种分布模型预测了卧龙保护区红腹角雉适宜栖息地分布,利用核密度估计方法分析了红腹角雉在卧龙保护区季节活动节律差异。该研究结果有利于深入了解卧龙保护区红腹角雉的栖息地分布、选择规律以及活动节律,可为保护区红腹角雉保护工作提供有针对性的保护策略。保护区红腹角雉适宜栖息地主要分布在皮条河两侧中低海拔区域,冬季红腹角雉趋于低海拔活动,接近居民聚集区,建议冬季严格管控人员上山活动,减少人为活动对红腹角雉产生干扰。同时红腹角雉为夜间树栖静息动物,易被捕猎,冬季应同时加强夜间巡查,避免偷盗猎等违法活动的发生。本文中采用的研究方法可为保护区其他雉类与不同雉类间生态位差异研究提供参考。红外相机技术经过数十年的发展与完善,目前已经成为野生动物调查与监测最有效方法之一,建议今后继续利用红外相机对保护区内的大中型哺乳动物以及地栖性鸟类进行长期监测,便于获取这些物种的种群、分布等长期动态变化的数据,以此来支持相关物种研究以及保护区的管理决策。
Study on Suitable Habitat and Activity Rhythm of Trgopan temminckii in Wolong National Nature Reserve
More Information-
摘要: 收集了卧龙国家级自然保护区2014—2019年红外相机调查中采集到的红腹角雉相关信息,使用物种分布模型(MaxEnt)预测红腹角雉在保护区内的适宜栖息地;采用核密度估计方法,分析了其在春夏秋冬四季活动节律差异。结果表明,在卧龙国家级自然保护区中,模型预测的红腹角雉潜在适宜栖息地面积为430.38 km2,占保护区总面积的21.52%,主要分布在保护区皮条河两侧中低海拔的针阔混交林中;在活动节律方面,红腹角雉在春季活动强度最高,秋季、夏季次之,冬季最低,红腹角雉在秋季的日活动节律与春季、夏季、冬季差异显著,而春季、夏季、冬季之间红腹角雉日活动节律无显著差异。本研究初步明确了红腹角雉在卧龙保护区内的潜在适宜栖息地分布以及春夏秋冬四季活动节律差异,为保护区红腹角雉乃至同域分布的其他雉类的保护与管理提供科学依据。Abstract: In this study, the relevant information about Trgopan temminckii collected by infrared camera survey in Wolong National Nature Reserve from 2014 to 2019 was collected, and species distribution model (MaxEnt) was used to predict the suitable habitat of Trgopan temminckii in the Nature Reserve. The difference of activity rhythm in spring, summer, autumn and winter was analyzed using kernel density estimation method. The results showed that in Wolong National Nature Reserve, the predicted suitable habitat for Trgopan temminckii was 430.38 km2, accounting for 21.52% of the total area of the reserve, which was mainly distributed in the coniferous and broad-leaved mixed forest at middle and low altitude on both sides of Pitiao River in the reserve. In terms of activity rhythm, the highest activity intensity of Trgopan temminckii was in spring followed by autumn and summer, and the lowest in winter. The daily activity rhythm of Trgopan temminckii in autumn was significantly different from that in spring, summer and winter, but there was no significant difference between spring, summer and winter. This study preliminarily identified the potential suitable habitat distribution of Trgopan temminckii in Wolong National Nature Reserve and the difference of activity rhythm in spring, summer, autumn and winter, which provided scientific basis for the conservation and management of Trgopan temminckii and other Phasianidaes distributed in the same area in the reserve.
-
图 1 研究区域地理位置及红腹角雉记录位点示意图
Fig. 1 Schematic map of the study area and the occurrence locations of Trgopan temminckii in the study area
图 3 红腹角雉在春季、夏季、秋季、冬季的活动节律曲线比较( 灰色为重叠区域)
Fig. 3 Comparison of daily activity rhythm of Trgopan temminckii in spring, summer, autumn and winter (The overlapping coefficient equals the area in grey)
表 1 红腹角雉适宜栖息地预测最终模型中使用的环境变量
Tab. 1 Environmental variables used in the final model of suitable habitat prediction of Trgopan temminckii
环境变量
Environmental variables分类与赋值
Classification and assignment原始分辨率
Original resolution来源
Source地形Topography 海拔 Elevation 连续变量Continuous variable 30 m 中国科学院计算机网络信息中心地理空间数据云平台Geospatial Data Gloud 坡度 Slope 0°–10° (1); 10°–20° (2); 20°–30° (3);30°–45° (4); > 45° (5) 坡向 Aspect 45°–225°(1) ; 225°–45°(2) 气候Climate 月均昼夜温Mean diurnal range (bio2) 连续变量Continuous variable 30″ WorldClim 最冷季度平均温度 Mean Temperature of Coldest Quarter (bio11) 连续变量Continuous variable 最湿月降水量 Precipitation of Wettest Month(bio13) 连续变量Continuous variable 最干月降水量Precipitation of Driest Monthbio (14) 连续变量Continuous variable 最暖季度降水量Precipitation of Warmest Quarter (bio18) 连续变量Continuous variable 植被Vegetation 归一化植被指数Normalized differenceVegetation index (NDVI) 连续变量Continuous variable 250 m Land Processes Distributed Active Archive Center 植被类型Land cover 高山流石滩 Alpine screes (1)
草甸草地 Meadow grassland (2)
高山灌丛 Alpine shrub (3)
针叶林 Coniferous forest (4)
针阔混交林Coniferous and broad-leaved mixed forest (5)
阔叶林 Broad-leaved forest (6)
其他 Others (7)500 m 人类干扰anthropogenic disturbance 人口密度Population density 连续变量Continuous variable 1 km 中科院资源环境数据云平台Resource and environment Data Gloud  下载: 导出CSV
下载: 导出CSV
-
[1] 李俊清, 大熊猫栖息地研究[M]. 北京: 高等教育出版社, 2012. [2] Reppert S M, Weaver D R. Coordination of circadian timing in mammals[J]. Nature, 2002, 418(6901): 935−941. doi: 10.1038/nature00965 [3] 张洪茂,胡锦矗. 川西北高原藏原羚夏季生境选择[J]. 四川动物,2002,21(1):12−15. doi: 10.3969/j.issn.1000-7083.2002.01.004 [4] 尚玉昌. 动物的行为节律[J]. 生物学通报,2006,41(10):8−10. doi: 10.3969/j.issn.0006-3193.2006.10.004 [5] Zhang Y B, Liu Y L, Fu J X, et al. Bridging the “Gap”in Systematic Conservation Planning[J]. Journal forNature Conservation, 2016, 31: 43−50. doi: 10.1016/j.jnc.2016.03.003 [6] Phillips S J, Anderson R P, Schapire R E. Maximum Entropy Modeling of Species Geographic Distributions[J]. Ecological Modelling, 2006, 190 (3): 231−259. [7] Zhang M G, Zhou Z K, Chen W Y, et al. Using Species Distribution Modeling to Improve Conservation and Land use Planning of Yunnan, China[J]. Biological Conservation, 2012, 153(x): 257−264. [8] Elith J, Graham C H, Anderson R P, et al. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data[J]. Ecography, 2006, 29(2): 129−151. doi: 10.1111/j.2006.0906-7590.04596.x [9] Phillips S J, Dudík M. Modeling of Species Distributions with Maxent: New Extensions and a Comprehensive Evaluation[J]. Ecography, 2008, 31(2): 161−175. doi: 10.1111/j.0906-7590.2008.5203.x [10] Phillips S J, Dudík M, Schapire R E. A maximum entropy approachto species distributio modeling[A]. Proceedings of the Twenty-First International Conference on Machine Learning[C]. 2004. [11] Banavar J R, Maritan A, Volkov I. Applications of the principle of maximum entropy: from physics to ecology[J]. Journal of Physics: Condensed Matter, 2010, 22(6): 3101. [12] 邢丁亮,郝占庆. 最大熵原理及其在生态学研究中的应用[J]. 生物多样性,2011,19(3):295−302. [13] 岳茂峰,冯莉,田兴山,等. 基于MaxEnt的入侵植物刺轴含羞草的适生分布区预测[J]. 生物安全学报,2013,22(3):173−180. doi: 10.3969/j.issn.2095-1787.2013.03.005 [14] 王茹琳,高晓清,王闫利,等. 基于MaxEnt的非洲橘硬蓟马在全球及中国的潜在分布区预测[J]. 中国农学通报,2014,30(28):315−320. doi: 10.11924/j.issn.1000-6850.2014-1016 [15] 李晟,王大军,肖治术,等. 红外相机技术在我国野生动物研究与保护中的应用前景[J]. 生物多样性,2014,22(6):865−695. [16] 李勤,邬建国,寇晓军,等. 相机陷阱在野生动物种群生态学中的应用[J]. 应用生态学报,2013,24:947−955. [17] O’Connell A F, Nichols J D, Karanth K U. Camera Trapsin Animal Ecology: Methods and Analyses [M]. NewYork: Springer, 2011. [18] 肖治术,李欣海,姜广顺. 红外相机技术在我国野生动物监测研究中的应用[J]. 生物多样性,2014,22:683−684. [19] Steenweg R, Hebblewhite M, Kays R, et al. Scaling-up camera traps: Monitoring the planet’s biodiv-ersity with networks of remote sensors[J]. Frontiers in Ecology and the Environment, 2017, 15: 26−34. doi: 10.1002/fee.1448 [20] Ridout MS, Linkie M. Estimating overlap of daily activity patterns from camera trap data[J]. Journal of Agricultural, Biological and Environmental Statistics, 2009, 14: 322−337. doi: 10.1198/jabes.2009.08038 [21] Oliveira-Santos LGR, Zucco CA, Agostinelli C. Using conditional circular kernel density functions to test hypo-theses on animal circadian activity[J]. Animal Behaviour, 2013, 85: 269−280. doi: 10.1016/j.anbehav.2012.09.033 [22] Rowcliffe JM, Kays R, Kranstauber B, et al. Quantifying levels of animal activity using camera trap data[J]. Methods in Ecology and Evolution, 2014, 5: 1170−1179. doi: 10.1111/2041-210X.12278 [23] 陈立军,束祖飞,肖治术. 应用红外相机数据研究动物活动节律—以广东车八岭保护区鸡形目鸟类为例[J]. 生物多样性,2019,27(3):266−272. doi: 10.17520/biods.2018178 [24] O’Connell AF, Nichols JD, Karanth KU. Camera Traps in Animal Ecology[M], Tokyo: Springer, 2011. [25] 郑光美, 王岐山. 中国濒危动物红皮书(鸟类)[M]. 北京: 科学出版社, 1998. [26] Zhang F C, Zheng G M, Zhou X P. The interspecific relationship between Tragopan temminckii and other birds-with the guild structure method[J]. Journal of Beijing Normal University: Natural Science, 1997, 33(3): 403−408. [27] 丛培昊. 红腹角雉的种群密度和繁殖行为研究[D]. 北京: 北京师范大学博士学位论文, 2007. [28] 崔鹏,邓文洪. 四川栗子坪自然保护区红腹角雉雌鸟和亚成体冬季集群行为研究[J]. 四川动物,2013,32(6):846−849. [29] 史海涛,郑光美. 红腹角雉的食性研究[J]. 动物学研究,1998,19(3):225−229. [30] 丛培昊,郑光美. 四川老君山地区红腹角雉的夜栖行为和夜栖地选择[J]. 生物多样性,2008,16(4):332−338. doi: 10.3321/j.issn:1005-0094.2008.04.003 [31] 崔鹏,康明江,邓文洪. 繁殖季节同域分布的红腹角雉和血雉的觅食生境选择[J]. 生物多样性,2008,16(2):143−149. doi: 10.3321/j.issn:1005-0094.2008.02.006 [32] 李湘涛. 红腹角雉的繁殖习性[J]. 动物学报,1987,33(1):99−100. [33] 李佳,刘芳,李迪强,等. 基于红外相机监测分析的红腹角雉日活动节律[J]. 林业科学,2017,53(7):170 −175. doi: 10.11707/j.1001-7488.20170717 [34] 刘小斌,韦伟,郑筱光,等. 红腹锦鸡和红腹角雉活动节律-基于红外相机监测数据[J]. 动物学杂志,2017,31(2):194−202. [35] 张福成,郑光美,周小平,等. 红腹角雉与其他鸟类种间关系研究-应用集团结构方法[J]. 北京师范大学学报 (自然科学版),1997,33(3):403−408. [36] 李仁贵,黄金燕,周世强,等. 卧龙大熊猫栖息地红腹角雉冬季生境选择的研究[J]. 四川林业科技,2011,32(2):55−59. doi: 10.3969/j.issn.1003-5508.2011.02.009 [37] 杨志松, 周材权, 何廷美, 等. 四川卧龙国家级自然保护区综合科学考察报告[M]. 北京: 中国林业出版社, 2019. [38] 王茹琳,李庆,封传红,等. 基于MaxEnt的西藏飞蝗在中国的适生区预测[J]. 生态学报,2017,37:8556− 8566. [39] 李娟. 青藏高原三江源地区雪豹(Panthera uncia)的生态学研究及保护[D]. 北京: 北京大学生命科学学院博士学位论文, 2012. [40] 李明富,李晟,王大军,等. 四川唐家河自然保护区扭角羚冬春季日活动模式研究[J]. 四川动物,2011,30(6):850−855. doi: 10.3969/j.issn.1000-7083.2011.06.031 [41] Schmid F, Schmidt R. Multivariate extensions of Spearman’s rho and related statistics[J]. Statistics and Probability Letters, 2006, 77: 407−416. [42] 易国栋,杨志杰,刘宇,等. 中华秋沙鸭越冬行为时间分配及日活动节律[J]. 生态学报,2010,30(8):2228−2234. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 545
- HTML全文浏览量: 201
- PDF下载量: 40
- 被引次数: 0








