






 下载:
下载:



-
杉木[(Cunninghamia lanceolata (Lamb.) Hook)]是我国特有的用材林树种,是南方各省区最重要的乡土造林树种之一,我国人民使用杉木的历史逾8 000年[1]。因此,对杉木进行持续、多目标的遗传改良,对于提高杉木遗传增益,培育多用途杉木品种具有重要意义。
杉木种子园是生产杉木良种的主要单元,也是进行杉木育种的重要载体,开展种子园人工控制授粉和遗传评价,建立世代清晰的高世代种子园和高改良轮次种子园是有效利用杉木遗传增益并保持种子园较高遗传基础的途径,为了充分发挥种子园的效能,育种工作者对杉木生殖发育、花粉散布规律进行了研究,对杉木大小孢子发育进程有较为完整的了解。而杉木在长期的进化过程中,为适用各地的气候,导致不同种源之间物候期的特征差异较大[2],四川是杉木分布的西部核心分布区,与东部和中部分区的杉木在物候上存在差异,而四川杉木减数分裂时期,小孢子叶球与小孢子发育的行为特征未见报道,杉木雄球花采集过早,小孢子尚处于发育阶段,干燥过程中容易失活死亡,采完了,则花粉开始散落,收集效果不佳。
以四川杉木种子园为研究对象,对同一小孢子叶球、同一小孢子叶上花粉的发育成熟状态进行了研究,并比较了不同基因型杉木花粉散布时期,以期为杉木杂交授粉中的花粉精准采收奠定基础。
-
供试杉木位于宜宾市高县月江国有林场的杉木2代种子园,属中亚热带湿润季风气候,年平均气温16.5 ℃,≥10 ℃的年积温5 288 ℃,年降水量1 214.4 mm,多集中在4—9月,年蒸发量1 034.3 mm,相对湿度85%,年平均日照时数1 064. 6 h,热量高,雨量丰沛,无霜期长,四季分明,下半年雨热同季。基地土壤主要由砖红色砂岩发育而成的山地黄壤和少量酸性紫色土构成。土层厚度60~80 cm,pH值4.60~5.05,土壤质地黏性,立地指数10~12,土层深厚、疏松、肥力中等[3]。
该杉木二代无性系种子园于1992年整地造林,1994年嫁接,由220个无性系采用顺序错位排列的方式进行配置。
-
2016年2月底,当最高气温达到9 ℃且持续7~10 d,在杉木种子园优树收集区内选择相邻的4棵不同基因型母株开始观察小孢子叶和小孢子叶球的发育过程,直至单株上的雄球花上不能飘散出花粉为止。
采集新鲜样品用醋酸洋红染色10 min后压片,在奥林巴斯BX-31显微镜下观察并拍照。
每个样品随机选择3个视野,某个时期的小孢子数量占比>80%则视此时期为取样小孢子叶球的发育时期。
2016—2019年,连续3年观察候选母株的散粉情况,记录散粉开始时期。
-
观察发现,小孢子叶球的发育程度与其主轴长度相关,当小孢子叶球发育至5 mm长、呈青色时,醋酸洋红染色镜检发现,中部小孢子囊中的小孢子进入四分体时期,可以清晰地发现,四分体的4个独立细胞排列成四面体型,萌发沟朝外(见图1A、B)。
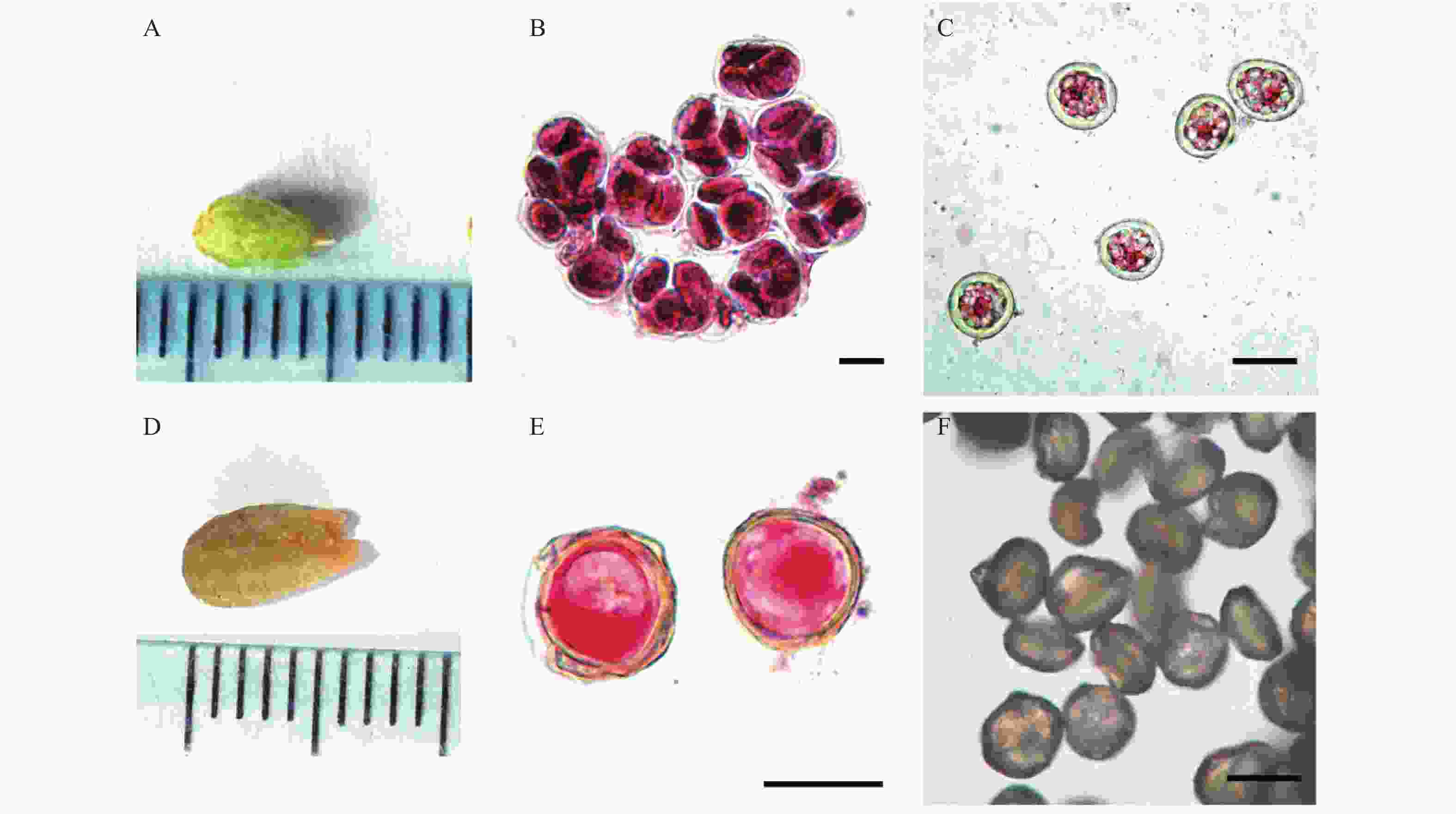
图 1 杉木小孢子发育进程(A,D:体式显微镜拍摄;B,C,E:醋酸洋红染色;F:花粉粒明场)
Figure 1. Development process of Chinese fir microspores (A, D: Stereomicroscope; B, C, E: Acetate magenta staining; F: Bright field of pollen grains).
同一个小孢子叶球上不同部位的小孢子叶发育进程基本同步,顶部小孢子叶的发育稍微比基部小孢子叶的发育快,但不会超过1个时期,即顶部小孢子的发育状态在基部小孢子中均能找到。
随着小孢子叶的伸长生长,小孢子叶颜色由青绿色转为淡黄色(见图1D),同时四分体解体为4个单核中央期的花粉粒,此时可见细胞核排列在细胞中央,细胞质稠密,没有液泡或只有小液泡出现(见图1C),此阶段的转化需要7~10 d,转化时长与天气状况紧密相关,晴天时间最短,阴雨天需要时间最长。采集此状态的小孢子叶球,放置在干燥温室内12~24 h,小孢子叶球干瘪,没有花粉释放。
当小孢子叶之间的鳞片逐步失水变黄,小孢子囊壁由青色变为淡黄色,花粉粒内形成了中央大液泡,细胞核被挤到细胞壁附近,细胞质所占比重少,小孢子发育至单核靠边期(见图1E)。小孢子从单核中央期发育至单核靠边期的时间为3~7 d。采集此状态的小孢子叶球,放置在干燥温室内12~24 h,可促进小孢子囊干燥并释放出花粉,镜检发现此类花粉的形态与自然状态下的释放的花粉完全一致,活力相当。
杉木花粉完全成熟时,花粉表面沉积物增多,醋酸洋红无法透过细胞壁进入花粉内。通过明场观察,可以发现花粉形似扁球体,花粉长40 μm左右,部分花粉的萌发沟上有一个乳头状突起,有的萌发沟内凹(见图1F)。单核靠边期小孢子发育成熟需要3~5 d。
当花粉发育成熟时,鳞片变为灰褐色(见图3A)、小孢子囊壁由黄色变成白色,小孢子叶球拉长至2~5 cm,小孢子间隔可达2~5 mm,阳光直射时,可见外翻的白色小孢子囊(见图2B,白色箭头),小孢子囊壁破裂后即可释放花粉。
来源于不同基因型母株上的雄球花发育规律一致,但进入散粉期的时间差3~7 d。2016—2019年散粉期观察发现同一基因型母株在不同年份散粉日期与气温相关,但不同年份各基因型进入散粉期的先后顺序一致。

图 2 同一簇上的小孢子叶球发育进程展示
Figure 2. Display of microstrobilus development process on the same cluster

图 3 即将进入散粉期的雄球花(A)和收集到的花粉(B)
Figure 3. Male cones in the pollinating period (A) and collected pollen (B)
-
同一个雄球花上的小孢子叶球表现为:位于中间部位的小孢子叶球发育最快,四周小孢子叶球发育较慢,而最外围小孢子叶球发育最慢(见图2A),但发育落差不会超过2个时期,从时间上分析,差6~12 d。
若雄球花所处环境太阳直射、通风条件好,则小孢子叶发育较快,有的小孢子叶完全成熟开始释放花粉(见图2B,白色箭头),而有的小孢子叶的小孢子囊排列紧密(见图2B,黑色箭头)收集处理此类雄球花的费效比较高,不建议采集。
因此,在收集雄球花时,应选择状态一致的雄球花,即小孢子叶球的颜色基本一致,鳞片为黄色、小孢子囊壁为淡黄色的雄球花,以及颜色比以上状态更深的雄球花均可。此时,大部分小孢子处于单核靠边期及花粉早期,温室处理能加快花粉的成熟(见图3B)。
干燥后的花粉,含水量低,可采用注射器喷施的方法进行杂交授粉。
-
采用人工控制授粉是建立子代测定林、测算遗传参数、筛选优良家系和单株的主要方法。本研究通过小孢子叶球外部形态与小孢子发育进程的对应关系,试验了各个发育阶段的雄球花干燥后收集花粉的比例,发掘适宜于温室干燥处理的雄球花采摘时期。
杉木生殖生物学研究丰富,从大小孢子发育[4]、花粉飘散规律到种子园花粉发育特征等各层面均有研究。本研究认为四川杉木四分体为四面体结构,与董金生[5]在江西的研究结论一致,而徐进[6]则认为杉木四分体主要是单平面分裂的方式,推测四分体状态与杉木种源有关系。
花粉飘散规律方面,张卓文对浙江、湖北多个种子园的花粉散发规律进行了研究[7],陈晓阳对贵州黎平杉木种子园花粉飞散进行了研究,提出半同胞子代测定的球果要从树冠上部采集[8]。陈波涛等研究了3个杉木种子园花粉释放和花粉密度,发现花粉飞散持续时间长短与天气状况紧密相关[9],本研究也得出了类似的结论。
不同地区杉木花芽开始分化的时期不同,如贵州黎平杉木5月下旬出现雄球花原基,而福建杉木6月下旬出现雄球花原基[10],湖北崇阳杉木5月中下旬开始雄球花分化[7]。而异地引种后杉木长时间栽培能改变其原有物候,与本地杉木的物候有一定程度的“趋同”现象[7]。杉木小孢子发生时期在单株之间的差异每年都一致[5],因此,人们所期望的种子园内完全随机交配很难发生,这与本研究连续3年的观察结果一致。
Development Characteristics of Chinese Fir Microspores and its Application in Cross Breeding
More Information-
摘要: 以四川杉木种子园母株为试验材料,观察春季雄球花和小孢子叶球外部形态发育与小孢子发育的对应关系,结果表明,同一个小孢子叶球上的小孢子叶发育基本同步;小孢子叶的颜色和长度与小孢子发育极其相关,小孢子叶球整体呈青色、长5 mm时,其内部小孢子基本处于四分体时期;当小孢子叶之间的鳞片失水变黄、小孢子囊壁变为淡黄色时,其内部小孢子基本处于单核靠边期,采集小孢子叶球为此状态的雄球花,在干燥温室中放置12~24 h后,能收获大量成熟的、活力旺盛的花粉。Abstract: Using the mother plant of Sichuan Cunninghamia lanceolata seed orchard as the test material, the corresponding relationship between the external morphological development of spring male cones, microstrobilus and the development of microspores were observed. The results showed that the development of microsporophyll on the same microstrobilus was basically synchronized. The color and length of microsporophyll were closely related to the development of microspores. When the whole microstrobilus was blue and up to 5 mm long, the internal microspores were basically in the tetrad stage. When the scales among the microsporophylls lost water and turned yellow and the wall of the microsporangia turned light yellow, the internal microspores were basically at the stage of mononuclear. After collecting the male cones with the microstrobiluses in this state and placing them in a dry greenhouse for 12—24 hours, a large number of mature and vigorous pollen could be harvested.
-
图 1 杉木小孢子发育进程(A,D:体式显微镜拍摄;B,C,E:醋酸洋红染色;F:花粉粒明场)
A:四分体时期小孢子叶球形态;B:四分体时期;C:单核中间期;D:单核中间期小孢子叶球形态;E:单核靠边期,左侧花粉可见中央大液泡;标尺=40 μm。
Fig. 1 Development process of Chinese fir microspores (A, D: Stereomicroscope; B, C, E: Acetate magenta staining; F: Bright field of pollen grains).
A: Morphology of microstrobilus at the tetrad stage; B: Tetrad stage; C: Mononuclear intermediate stage; D: Morphology of microstrobilus in the mononuclear intermediate stage; E: Mononuclear at the side stage, left pollen has large vacuoles; F: Mature pollen. Ruler=40 μm.
图 2 同一簇上的小孢子叶球发育进程展示
Fig. 2 Display of microstrobilus development process on the same cluster
-
[1] 林幼丹,张晨曦. 杉木在中国的栽培历史简述[J]. 自然辩证法通讯,2007,29(1):79−82. doi: 10.3969/j.issn.1000-0763.2007.01.013 [2] 傅远志. 杉木花粉形态的研究[J]. 浙江林业科技,1994,14(6):9−12. [3] 杨平,陈炙,黄振,等. 月江杉木2代种子园优树选择研究[J]. 四川林业科技,2016(3):20−24. [4] 李齐,陈晓阳,张天琪. 贵州黎平杉木花芽分化的观察[J]. 北京林业大学学报,1996(2):100−102. [5] 董金生. 杉木小孢子发生与花粉发育的研究[J]. 江西农业大学学报,1985(3):29−34. [6] 徐进,施季森,杨立伟,等. 杉木花粉母细胞减数分裂的细胞学特性及异常现象的观察[J]. 林业科学,2007(11):32−36. doi: 10.3321/j.issn:1001-7488.2007.11.006 [7] 张卓文. 杉木生殖生物学特性研究[D]. 华中农业大学, 2005. [8] 陈晓阳,松丙龙,杨祖华,等. 杉木种子园内花粉飞散的观测与分析[J]. 北京林业大学学报,1991(2):48−53. [9] 陈波涛,王欣,张贵云,等. 杉木种子园花粉管理技术研究[J]. 贵州林业科技,1998(1):1−8. [10] 俞新妥,陈存及,白育玲,等. 杉木花芽分化的观察[J]. 林业科学,1981(1):46−49. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 430
- HTML全文浏览量: 124
- PDF下载量: 18
- 被引次数: 0








